Независимата наука за ефекта на безжичната радиация върху човешкото здраве
Независима наука за ефекта на безжичната радиация върху човешкото здраве

U
Има повече от 1000 научни изследвания, проведени от независими изследователи от цял свят относно биологичните ефекти на радиочестотната радиация. Тук представяме някои от най-новите.
I. ефекти върху феталното развитие и новороденото
Излагането на майката на електромагнитни полета преди и по време на бременност е свързано с риск от проблеми с говора при потомството. Zarei, S., et al. Вестник по биомедицинска физика и инженерство 9 (1): 61-68 (2019).
Излагането на майката на електромагнитни полета преди и по време на бременност е свързано с риск от речеви проблеми при потомството
Принадлежности- PMID: 30881935
- PMCID: PMC6409372
Резюме
Предистория: Бързият напредък в технологиите, особено в областта на телекомуникациите, доведе до извънредни нива на излагане на майките на радиочестотни електромагнитни полета (RF-EMF) преди или по време на бременност.
Цел: Основната цел на това проучване беше да се отговори на този въпрос дали излагането на жените на често срещани източници на RF-EMF, преди или по време на бременност, е свързано с речеви проблеми при потомството.
Материали и методи: В това проучване бяха интервюирани майки на 110 деца от три до седем години с речеви проблеми и 75 здрави деца (контролна група). Тези майки бяха попитани дали са били изложени на различни източници на ЕМП, като мобилни телефони, мобилни базови станции, Wi-Fi, безжични телефони, лаптопи и електропроводи. Тестът с хи квадрат беше използван за анализ на разликите, наблюдавани между контролната и изложената група.
Резултати: Установени са статистически значими връзки между използването на безжичен телефон и проблеми с речта на потомството както преди бременността, така и по време на бременност при майки (P = 0,005 и P = 0,014, съответно). Въпреки това, поради високата честота на използване на мобилни телефони и в двете групи, това проучване не успя да покаже никаква връзка между използването на мобилни телефони и речевите проблеми при потомството. Освен това се наблюдават значителни асоциации между живота в близост до електропроводи и речевите проблеми отново както преди бременността, така и по време на бременност при майки (P = 0,003 и P = 0,002, съответно). Излагането на други източници на нейонизиращо лъчение обаче не е свързано с речеви проблеми. Освен това излагането на йонизиращо лъчение (напр. Рентгенография преди и по време на бременност) не е свързано с появата на речеви проблеми.
Заключение: Въпреки че това проучване има някои ограничения, то ни води до този извод, че по-високите от всякога нива на майчина експозиция на електромагнитни полета могат да бъдат свързани с речеви проблеми на потомството.
Ключови думи: Електромагнитни полета; Излагане; Йонизиращо лъчение; Нейонизиращо лъчение; Бременност; Речев проблем.
Подобни статии
- Предизвикателен въпрос в етиологията на речевите проблеми: Ефектът от излагането на майката на електромагнитни полета върху речевите проблеми при потомството.J Biomed Phys Eng. 2015 г. 1 септември; 5 (3): 151-4. eCollection 2015 септември.PMID: 26396971 Безплатна статия от PMC.
- Радиочестотни електромагнитни полета, време на екрана и емоционални и поведенчески проблеми при 5-годишни деца.Int J Hyg Environment Health. 2019 март; 222 (2): 188-194. doi: 10.1016 / j.ijheh.2018.09.006. Epub 2018 9 октомври.PMID: 30314943
- Проучване на ефектите от излагането на 900 MHz радиочестотно излъчване, излъчвано от GSM мобилен телефон, върху модела на мускулни контракции в животински модел.J Biomed Phys Eng. 2015 г., 1 септември; 5 (3): 121-32. eCollection 2015 септември.PMID: 26396968 Безплатна статия от PMC.
- Насоки EUROPAEM EMF 2016 за профилактика, диагностика и лечение на здравни проблеми и заболявания, свързани с ЕМП.Rev Environment Health. 2016 г. 1 септември; 31 (3): 363-97. doi: 10.1515 / reveh-2016-0011.PMID: 27454111 Преглед.
- Биологични ефекти от експозицията на електромагнитно поле и обществените стандарти за експозиция.Biomed Pharmacother. 2008 февруари; 62 (2): 104-9. doi: 10.1016 / j.biopha.2007.12.004. Epub 2007 31 дек.PMID: 18242044 Преглед.
Препратки
- Peart KN [Интернет] Използването на мобилен телефон по време на бременност може да причини поведенчески разстройства при потомството. [цитирано на 15 март 2012 г.] Достъпно от: [https://news.yale.edu/2012/03/15/cell-phone-use-pregnancy-may-cause-behavioral-disorders-offspring. ]
- Zarei S, Mortazavi SM, Mehdizadeh AR, Jalalipour M, Borzou S, Taeb S, et al. Предизвикателен въпрос в етиологията на речевите проблеми: Ефектът от излагането на майката на електромагнитни полета върху речевите проблеми при потомството. J Biomed Phys Eng. 2015; 5: 151–4. [Безплатна статия от PMC] - PMC - PubMed
- Mokarram P, Sheikhi M, Mortazavi SMJ, Saeb S, Shokrpour N. Ефект от излагане на 900 MHz GSM радиочестотна радиация на мобилен телефон върху състоянието на метилиране на естрогенния рецептор в клетките на дебелото черво на мъжки Sprague Dawley плъхове. J Biomed Phys Eng. 2017; 7: 79–86. [Безплатна статия от PMC] - PMC - PubMed
Пренатална експозиция на изключително нискочестотно магнитно поле и неговото въздействие върху феталния растеж. Ren, Y., et al. Екологично здраве (2019).
Пренатално излагане на изключително нискочестотно магнитно поле и неговото въздействие върху растежа на плода
Принадлежности- PMID: 30635061
- PMCID: PMC6329146
- DOI: 10.1186 / s12940-019-0447-9
Резюме
Цел: Изследванията върху ефекта от пренаталното излагане на магнитно поле (MF) върху растежа на плода са неубедителни и са обект на някои методологични ограничения, особено при измерване на експозицията на MF. Настоящото проучване има за цел да изследва връзката между експозицията на майки с изключително ниска честота (ELF-MF) по време на бременност и растежа на плода при потомството.
Методи: Общо 128 бременни жени са били наети на работа през третия триместър и са били помолени да носят измервателен уред EMDEX Lite в продължение на 24 часа, за да уловят ежедневната експозиция на ELF-MF. Средно претеглените във времето (TWA), P50 и P75 лични 24-часови измервания са използвани за оценка на пренаталната експозиция на ELF-MF. Медианите на тези измервания са използвани като гранични точки на висока и ниска пренатална експозиция на ELF-MF. Растежът на плода се измерва чрез теглото при раждане на бебето, дебелината на кожните гънки на трицепсите, корема и гърба и обиколката на главата, горната част на ръката и корема. Тези мерки бяха проведени в рамките на 24 часа след раждането. Използва се генерализиран линеен модел, за да се изследва връзката между нивото на ELF-MF при майката и индексите на растежа на плода след коригиране на потенциалните смутители.
Резултати: В сравнение с момичетата с по-ниска пренатална експозиция на ELF-MF, момичетата с по-висока експозиция имат по-ниско тегло при раждане, по-тънка кожна гънка на трицепсите, корема и гърба и по-малка обиколка на главата, горната част на ръката и корема във всичките три ELF-MF матрици. Разликите са статистически значими за теглото при раждане и повечето други измервания на растежа (P <0,05). Тези мерки не са имали значителна разлика между по-висока и по-ниска пренатална експозиция на ELF-MF при момчетата, с изключение на дебелината на кожата на гърба.
Заключение: Пренаталното излагане на по-високи нива на ELF-MF е свързано с намален растеж на плода при момичета, но не и при момчета.
Ключови думи: Кохортно проучване; Растеж на плода; Магнитно поле; Бременност.
Изявление за конфликт на интереси
Етично одобрение и съгласие за участие
Изследването е одобрено от комитета за етичен преглед на Шанхайския институт за планирани изследвания на родителството (SIPPR). Изследването е проведено съгласно Декларацията от Хелзинки. Жените, участващи в проучването, са предоставили писмено информирано съгласие преди провеждането на проучването.
Съгласие за публикуване
Не е приложимо.
Конкуриращи се интереси
Авторите заявяват, че нямат конкуриращи се интереси.
Бележка на издателя
Springer Nature остава неутрален по отношение на юрисдикционните претенции в публикувани карти и институционални принадлежности.
Подобни статии
- Излагане на майката на изключително нискочестотни магнитни полета: Асоциация с времето към бременността и растежа на плода.Environment Int. 2016 септември; 94: 620-625. doi: 10.1016 / j.envint.2016.06.027. Epub 2016 7 юли.PMID: 27395334
- Излагане на бисфенол А и фталати по време на бременност и ултразвукови мерки за фетален растеж в кохортата INMA-Sabadell.Environmental Health Perspect. 2016 април; 124 (4): 521-8. doi: 10.1289 / ehp.1409190. Epub 2015 21 юли.PMID: 26196298 Безплатна статия от PMC.
- Сравнение на монитори за лична експозиция с изключително ниска честота (ELF) с магнитно поле.J Expo Anal Environment Epidemiol. 2002 г. януари-февруари; 12 (1): 1-8. doi: 10.1038 / sj.jea.7500194.PMID: 11859428
- Размер на тялото на потомството и метаболитен профил - ефекти от намесата в начина на живот при затлъстели бременни жени.Dan Med J. 2014 юли; 61 (7): B4893.PMID: 25123127 Преглед.
- Изключително нискочестотните магнитни полета предизвикват апоптоза на сперматогенни зародишни клетки: възможен механизъм.Biomed Res Int. 2014; 2014: 567183. doi: 10.1155 / 2014/567183. Epub 2014 15 юни.PMID: 25025060 Безплатна статия от PMC. Преглед.
Препратки
- Файнман. R: Лекциите на Feynman по физика Vol II: Addison Wesley Longman. ISBN 978-0-201-02115-8; 1970 г.
- Carlo V, Bellieni I. Bioinitiative Report, раздел 19, фетални и неонатални ефекти на ЕМП. 2012 г.
- Behari J. BioInitiative Report, раздел 18, Ефекти на експозиция на електромагнитно поле (ELF-EMF и RFR) върху плодовитостта и възпроизводството. 2012 г.
- Li DK, Odouli R, Wi S, Janevic T, Golditch I, Bracken TD, Senior R, Rankin R, Iriye R. Популационно проспективно кохортно проучване на лична експозиция на магнитни полета по време на бременност и риск от спонтанен аборт. Епидемиология. 2002; 13 (1): 9–20. doi: 10.1097 / 00001648-200201000-00004. - DOI - PubMed
Ефектите на радиочестотното излъчване върху теглото, дължината и тъканите на плода на мишките. Alimohammadi, I., et al. Данни накратко 19: 2189-2194 (2018).
Ефекти от пренаталното излагане на WiFi сигнал (2,45 GHz) върху постнаталното развитие и поведение при плъхове: Влияние на майчините ограничения. Othman, H., et al. Изследвания на поведенческия мозък 326: 291-301 (2017).
Ефекти от пренаталното излагане на WIFI сигнал (2,45 GHz) върху постнаталното развитие и поведение при плъхове: Влияние на сдържаността на майката
Принадлежности- PMID: 28288806
- DOI: 10.1016 / j.bbr.2017.03.011
Резюме
Настоящото проучване беше проведено, за да се изследва потенциалното комбинирано влияние на стрес от майката и излагане на 2,45 GHz WiFi сигнал върху постнаталното развитие и поведението при потомството на изложени плъхове. 24 бременни плъхове албиноси Wistar бяха разпределени на случаен принцип в четири групи: контролни, изложени на WiFi, задържани и двете изложени на WiFi и задържани групи. Всяко излагане и ограничаване на WiFi се случва 2 часа на ден по време на бременността до раждането. Кученцата бяха оценени за физическо развитие и невромоторно съзряване. Нещо повече, тест с повишен плюс лабиринт, активност в открито поле и стационарен тест на лъча също бяха определени съответно на 28, 30 и 31 ден след раждането. След поведенчески тестове плъховете се упояват и мозъците им се отстраняват за биохимичен анализ. Основните ни открития не показват вредно въздействие върху напредъка на бременността и резултатите при раждането във всички групи. Впоследствие WiFi и сдържаност, сами по себе си и главно в съгласие променят физическото развитие на малките с малки разлики между половете. Поведенчески, гестационното WiFi облъчване, сдържаност и особено свързаното лечение повлияват невромоторното съзряване главно при мъжкото потомство. В зряла възраст забелязахме тревожност, двигателен дефицит и нарушение на експлораторното поведение при мъжкото потомство, изложено на WiFi облъчване и ограничение, и при женското потомство, подложено на три лечения. Биохимичното изследване показа, че и трите лечения са произвели глобален оксидативен стрес в мозъка и на двата пола. Що се отнася до серумната биохимия, нивата на фосфор, магнезий, глюкоза, триглицериди и калций бяха нарушени. Взети заедно,
Ключови думи: 2,45 GHz WiFi сигнал; Поведение; Невроразвитие; Потомство; Оксидативен стрес; Пренатална; Сдържаност; Серумна биохимия.
Copyright © 2017 Elsevier BV Всички права запазени.
Подобни статии
- Постнатално развитие и ефекти на поведението при вътреутробно излагане на плъхове на радиочестотни вълни, излъчвани от конвенционални WiFi устройства.Environment Toxicol Pharmacol. 2017 юни; 52: 239-247. doi: 10.1016 / j.etap.2017.04.016. Epub 2017 22 април.PMID: 28458069
- Едновременно излагане на алуминий и стрес по време на бременност при плъхове: Ефекти върху постнаталното развитие и поведението на потомството.Невротоксикол Тератол. 2005 юли-август; 27 (4): 565-74. doi: 10.1016 / j.ntt.2005.06.014.PMID: 16024221
- Ефекти на повтарящ се стрес и експозиция на WiFi сигнал върху поведението и оксидативен стрес при плъхове.Metab Brain Dis. 2017 октомври; 32 (5): 1459-1469. doi: 10.1007 / s11011-017-0016-2. Epub 2017 27 април.PMID: 28451780
- Механизми на клетъчния стрес на пренаталния майчин стрес: Фактори на топлинен шок и оксидативен стрес.Neurosci Lett. 2019 г., 14 септември; 709: 134368. doi: 10.1016 / j.neulet.2019.134368. Epub 2019 9 юли.PMID: 31299286 Безплатна статия от PMC. Преглед.
- Поведенчески аномалии, медиирани от мъже.Mutat Res. 1990 април; 229 (2): 213-29. doi: 10.1016 / 0027-5107 (90) 90095-l.PMID: 2181296 Преглед.
Цитирано от 3 статии
- Асоциация между излагането на майката на нейонизиращо лъчение на магнитно поле по време на бременност и риск от разстройство с дефицит на внимание / хиперактивност при потомство в надлъжна кохорта при раждане.JAMA Netw Open. 2020 г. 2 март; 3 (3): e201417. doi: 10.1001 / jamanetworkopen.2020.1417.PMID: 32207831 Безплатна статия от PMC.
- Диетични профили, хранителен биохимичен статус и разстройство с дефицит на внимание / хиперактивност: Анализ на пътя за проучване на случай-контрол.J Clin Med. 2019 май 18; 8 (5): 709. doi: 10.3390 / jcm8050709.PMID: 31109092 Безплатна статия от PMC.
- Последен напредък в ефектите на микровълновото лъчение върху мозъка.Mil Med Res. 2017 г., 21 септември; 4 (1): 29. doi: 10.1186 / s40779-017-0139-0.PMID: 29502514 Безплатна статия от PMC. Преглед.
Излагане на нейонизиращо лъчение на магнитно поле и риск от спонтанен аборт: Проспективно кохортно проучване. Li, De-Kun, et al. Научни доклади (2017).
Излагане на нейонизиращо лъчение на магнитно поле и риск от спонтанен аборт: Проспективно кохортно проучване
Научни доклади 7 , Номер на артикул : 17541 ( 2017 )
Резюме
Нейонизиращото лъчение на магнитното поле (MF) е широко разпространено и всички са изложени до известна степен. Това проспективно кохортно проучване на 913 бременни жени изследва връзката между високата експозиция на МФ и риска от спонтанен аборт. За изследване на асоциацията е използвана регресия на Кокс (пропорционални опасности). След контролиране на множество други фактори, жените, които са били изложени на по-високи нива на МФ, са имали 2,72 пъти риск от спонтанен аборт (коефициент на риск = 2,72, 95% ДИ: 1,42–5,19) от тези с по-ниска експозиция на МФ. Постоянно се наблюдава повишен риск от спонтанен аборт, свързан с висока MF, независимо от източниците на висока MF. Асоциацията е много по-силна, ако MF се измерва в типичен ден от бременността на участниците. Констатацията също демонстрира, че точното измерване на експозицията на MF е жизненоважно за изследване на ефектите върху MF върху здравето.
Въведение
Нейонизиращото лъчение на магнитното поле (MF) е повсеместно излагане на околната среда и сериозно заплашващо предизвикателство за общественото здраве. MF се излъчват както от традиционни източници, които генерират нискочестотни MF (напр. Електропроводи, уреди, трансформатори и т.н.), така и от възникващи източници, които генерират MF с по-висока честота (напр. Безжични мрежи, интелигентни мрежови измервателни уреди, клетъчни кули, безжични устройства като като мобилни телефони и др.). Понастоящем хората са широко изложени на MF с постоянно нарастваща интензивност, поради разпространението на апарати, генериращи MF.
Рязкото нарастване на експозицията на МФ поднови опасенията относно потенциалните ефекти върху здравето от това невидимо, предизвикано от човека излагане на околната среда. Неотдавнашен многогодишен проект на NIEHS, проведен от Националната токсикологична програма (NTP), разкри повишен риск от рак, свързан с излагане на MF на нейонизиращо лъчение 1 , 2 . По-конкретно, проучването NTP установи, че рискът от рак, дължащ се на експозиция на MF, наблюдавана при техните експериментални животни, съответства на типовете ракови клетки, за които се съобщава в предишни епидемиологични проучвания при човешки популации 1. Това откритие затруднява продължаването на отхвърлянето на възможните биологични ефекти от експозицията на MF. Подобно уволнение би могло да бъде особено обезпокоително предвид голямото разпространение на излагането на хора (като почти всички са били изложени на MF нейонизиращо лъчение до известна степен). Това включва уязвими групи от населението като бременни жени и малки деца. Международната агенция за изследване на рака (IARC) класифицира MF като възможен канцероген 3 , 4 .
Спонтанният аборт е един от потенциалните неблагоприятни последици за здравето, които са чувствителни към експозицията на MF, а също и крайна точка, която СЗО препоръчва да бъде допълнително проучена в контекста на MF ефектите върху здравето 5 . През годините няколко наблюдателни проучвания при човешки популации предполагат възможна връзка между експозицията на МФ по време на бременност и повишения риск от спонтанен аборт 6 , 7 , 8 , 9 , 10 , 11, включително две проучвания, публикувани през 2002 г., които повишават обществената информираност за такава асоциация 12 , 13. В допълнение, едно проучване изследва човешки ембрионални тъкани, за да се оцени връзката между експозицията на ЕМП и ембрионалния растеж и наблюдава повишен риск от нарушен растеж на ембрионални пъпки и апоптоза, свързани с излагане на по-високо ниво на MF 14 , предоставяйки някои преки доказателства за неблагоприятно биологично въздействие Излагане на ЕМП върху ембрионалното развитие.
Независимо от това, връзката между експозицията на МФ и риска от спонтанен аборт остава до голяма степен неизвестна и пренебрегвана. Проведохме това проспективно кохортно проучване сред голяма популация бременни жени, за да проучим допълнително дали излагането на MF нейонизиращо лъчение по време на бременност увеличава риска от спонтанен аборт.
Материали и методи
Това проспективно кохортно проучване беше одобрено от Институционалния съвет за преглед на Kaiser Permanente в Северна Калифорния (KPNC) и проведено сред бременните членове на KPNC в района на залива на Сан Франциско, всички от които предоставиха информирано съгласие. Изследването е проведено в съответствие с всички съответни насоки и разпоредби. KPNC е интегрирана система за предоставяне на здравни грижи, чиито членове съставляват 28–30% от населението в водосборния басейн и последователно е доказано, че са представителни за основното население 15 , 16 .
Проучване на популацията
Всички бременни жени на възраст над 18 години и пребиваващи в участващите окръзи на залива са идентифицирани чрез лабораторната база данни на KPNC електронна медицинска карта (EMR) въз основа на положителни тестове за бременност. В KPNC всички жени, за които се подозира, че са бременни, рутинно са били помолени да се направи тест за бременност в център за KPNC. Флаери, информиращи жените за проучването, бяха публикувани в участващите съоръжения и дадени на жените по време на теста за бременност. Като се има предвид, че спонтанен аборт може да се случи много рано по време на бременността, набирането на бременни жени възможно най-рано по време на бременността е от решаващо значение за осигуряване на възможно най-пълно установяване на спонтанен аборт. Нашата идентификация на бременни жени чрез положителни лабораторни тестове за бременност осигури ранно набиране. За да определите дали повтарящи се аборти на жената, индикация за по-висока податливост на спонтанен аборт, увеличава нейната уязвимост към експозиция на МФ, ние взехме проби от жени с два или повече предишни спонтанни аборта. Идентифицираните бременни жени се свързват с обучен рекрутер / интервюиращ, за да се определи тяхната допустимост и желание за участие в проучването. Тези, които са заявили намерението си да пренесат бременността до края и чиято гестационна възраст при идентификация е била по-малка от 10 завършени седмици (все още в риск от спонтанен аборт), са поканени да участват в проучването. Сред 1627 допустими бременни жени, 1054 се съгласиха да участват в проучването. Идентифицираните бременни жени се свързват с обучен рекрутер / интервюиращ, за да се определи тяхната допустимост и желание да участват в проучването. Тези, които са заявили намерението си да пренесат бременността до края и чиято гестационна възраст при идентификация е била по-малка от 10 завършени седмици (все още в риск от спонтанен аборт), са поканени да участват в проучването. Сред 1627 допустими бременни жени, 1054 се съгласиха да участват в проучването. Идентифицираните бременни жени се свързват с обучен рекрутер / интервюиращ, за да се определи тяхната допустимост и желание да участват в проучването. Тези, които са заявили намерението си да пренесат бременността до края и чиято гестационна възраст при идентификация е била по-малка от 10 завършени седмици (все още в риск от спонтанен аборт), са поканени да участват в проучването. Сред 1627 допустими бременни жени, 1054 се съгласиха да участват в проучването.
Измерване на експозицията на магнитно поле по време на бременност
Всички участващи бременни жени бяха помолени да носят измервателен уред EMDEX Lite (Enertech Consultants Inc.) за 24 часа по време на бременността. Измервателният уред EMDEX Lite е специално проектиран за измерване на MF, който се измерва в милигауси (mG).
За да осигурим по-добро представяне на експозицията на МФ по време на бременност и да приложим знанията, получени от предишното проучване 12 , ние проектирахме измерването на МФ да се извършва в обикновен ден (ден, отразяващ типичния модел на участниците в работата и развлеченията по време на бременност). В случай, че ежедневните дейности на даден участник може да са били променени от първоначално планираните, ние също проверихме с участниците в края на периода на измерване дали денят на измерване наистина е типичен ден за тяхната бременност. Ако не, денят на измерване е класифициран като нетипичен.
Измервателят EMDEX Lite е използван за измерване на нивата на експозиция на MF чрез участващи бременни жени от всички източници на излъчване. Участниците също бяха помолени да водят дневник през 24-часовия период на измерване, за да позволят на изследователите (1) да идентифицират местата на ежедневните дейности (у дома, у дома в леглото, в движение, на работа и други), (2) проверете дали дейностите отразяват обичайния ден и (3) проверете дали местата и дейностите са свързани с висока експозиция на МФ.
Данните за MF заедно с дневника на участниците за дейностите в деня на измерване бяха изследвани за контрол на качеството, включително последователност и потенциални грешки. Изключихме 31 субекта, които не са успели да носят уреда според указанията. Също така изключихме 107 субекта, които са имали непълни (<90% от техните 24-часови измервания) MF данни за измерване. Тези изключения са направени без знание за резултатите от бременността на субектите.
Предишни проучвания са установили, че най-високите нива на MF, които бременните жени срещат, са най-подходящи за риска от спонтанен аборт 12 , 13 , което показва възможен прагов ефект при дадено ниво на MF, над който ембрионите в развитието могат да престанат да бъдат жизнеспособни. По този начин, това проучване се фокусира върху високите нива на експозиция на MF. Използвахме 99 -ия процентил от измерванията на MF през 24-часовия период, за да класифицираме нивото на експозиция, балансирайки между необходимостта да се изследва възможно най-високото ниво на MF и в същото време да избягваме използването на по-малко стабилни индекси (напр. Максимална експозиция ниво).
За да отразяваме по-точно истинската експозиция на МФ по време на бременност, положихме значителни усилия да отделим тези участници, чиито измервания бяха проведени по време на типичен ден от бременността им, от тези, чиито измервания не бяха проведени в обикновен ден. Измерванията, получени в типичен ден , вероятно са по-представителни за експозицията на МФ по време на бременност, докато измерванията, получени в нетипиченден са по-обект на погрешно представяне на истинското ниво на експозиция на МФ по време на бременност, което води до погрешно класифициране на участниците в неправилни категории на експозиция на МФ. Подобна погрешна класификация обикновено намалява способността на учените да откриват основна асоциация. Както се вижда от предишно проучване, измервания, проведени на един типичен ден показаха по-силна връзка между излагането на МФ и спонтанен аборт на риска, а измервания, проведени върху една нетипична ден показаха почти никаква връзка се дължи на неправилно класифициране на участниците в МФ категории на експозиция 12 .
Измерване на спонтанен аборт
Използвайки KPNC EMR данни, успяхме да идентифицираме бременността на участниците веднага след положителен тест за бременност, като по този начин започнахме проследяване на по-ранна гестационна възраст от първото пренатално посещение, най-ранното време, в което повечето други проучвания са успели да идентифицират бременна Жени. Това ранно проследяване ни позволи да установим ранни спонтанни аборти, които повечето други проучвания биха пропуснали, което го прави важна сила на това проучване.
Всички участници бяха проследени за резултатите от тяхната бременност от момента на техния положителен тест за бременност до края на бременността. В случай на спонтанен аборт това по дефиниция е преди 20 завършени гестационни седмици. Ние установихме резултатите от бременността чрез базата данни на KPNC EMR. За участниците, чиито резултати не бяха налични в EMR, ние се свързахме директно с тях. Успяхме да идентифицираме резултатите от бременността за всички участници, с изключение на една, която се е преместила извън района, поради което тя беше изключена от по-нататъшен анализ.
Лично интервю
Проведено е лично интервю с всички участници, за да се установи обширна информация за потенциалните смущаващи фактори, включително история на бременността и рискови фактори за спонтанен аборт. Предишни проучвания показват, че нивото на експозиция на МФ рядко е свързано с общи социално-демографски характеристики и рискови фактори 12 , 17 , 18 ; по този начин броят на потенциалните смутители в това проучване беше малък. Въпреки това все пак събрахме много фактори за изследване, за да осигурим задълбочен контрол на объркващите. Двама участници не успяха да попълнят интервюто, поради което бяха изключени от анализите.
Проектът на потенциалното проучване също така гарантира, че личното интервю е заслепено от експозицията на MF както за интервюиращите, така и за участниците, тъй като измерването на EMF е проведено след интервюто. Този дизайн на проучването подобрява качеството на резултатите от проучването.
Статистически анализ
Използвахме регресионния модел на Cox Proportional Hazards, с приспособяване за ляво отрязване, за да изследваме връзката между нивото на експозиция на MF и спонтанен аборт За определяне на големината и значимостта на асоциациите са използвани съотношения на риск с 95% доверителни интервали. Лявото съкращаване възниква, когато участниците в изследването влязат в наблюдение в определен момент от време (т.е. гестационна възраст при влизане в кохорта) след времето на произход, зачеването. Участниците са били проследявани до (а) спонтанен аборт, (б) край на бременността поради други резултати (напр. Извънматочна бременност), в който момент са били цензурирани или (в) 20 гестационни седмици за участници, които са останали бременни по това време .
Изследвахме объркващите, използвайки критерия за промяна в оценката, включително конфудера, ако съотношението на риска от спонтанен аборт (HR) за MF се промени с 10% или повече. Въпреки че повечето изследвани фактори не бяха объркващи поради липса на връзка с експозицията на МФ, въпреки това включихме в модела общоизвестни рискови фактори за спонтанен аборт и социално-демографски характеристики.
Като се има предвид предишното откритие, че силата на връзката между MF и спонтанен аборт варира в зависимост от това дали измерванията на MF са взети в типичен или нетипичен ден 12 , първо проведохме анализи отделно по тип ден. Предишното откритие беше потвърдено в настоящото проучване и поради това проведохме останалите анализи само сред онези, чиято експозиция на MF беше измерена в типичен ден от бременността им.
Тъй като препробовахме тези с множество предишни спонтанни аборти, първо направихме стратифициран анализ от тези с и без множество предишни спонтанни аборти, за да определим дали MF асоциацията с риск от спонтанен аборт се различава между тези две групи. След като беше установено, че наблюдаваните асоциации са до голяма степен сходни, включихме всички участници в анализите и коригирахме за предварителен спонтанен аборт във всички модели.
Общо 913 субекта с валидни MF измервания и резултати от бременността бяха включени в крайния анализ.
Статистически анализи бяха проведени с помощта на SAS 9.3.
Резултати
Таблица 1 представя описанието и характеристиките на участниците въз основа на техните нива на експозиция на MF (високо спрямо ниско). Групата с ниска експозиция на MF се състои от жени, чийто 99 -ти процентил от нивата на експозиция на MF е в най-ниския квартил (<2,5 mG), докато тези в горните три квартила са класифицирани в групата с висока експозиция на MF. Няма забележими връзки или последователни модели между нивото на експозиция на МФ и повечето от изследваните фактори, включително рискови фактори за спонтанен аборт (Таблица 1 ).
Таблица 1 Характеристики на изследваната популация по дневно ниво на експозиция на магнитно поле (най-ниски или по-високи квартили от MF 99 -ти процентил) След корекция за възрастта на майката, расата, образованието, тютюнопушенето по време на бременност и предишния спонтанен аборт, като цяло, бременните жени, които са имали по-висока експозиция на МФ по време на бременност (по-високи 3 квартила) са имали 48% по-голям риск от спонтанен аборт, отколкото жените, които са имали по-ниска експозиция на МФ ( в най-ниския квартил): коригирана HR = 1,48, 95% доверителен интервал (CI): 1,03–2,14 (Таблица 2 ). Забележително е, че в съответствие с констатацията в предишно проучване 12 , наблюдаваната асоциация е много по-силна сред участниците, чиято експозиция на MF е измерена в типичен ден от бременността (aHR = 2.72, 1.42–5.19). За разлика от това, не е наблюдавана връзка между тези, чиято MF е измерена в нетипичен ден (Таблица 2). По този начин, следващите анализи бяха ограничени до тези, чиято MF беше измерена в типичен ден от бременността им.
Таблица 2 Излагане на силни магнитни полета (МФ) по време на бременност и риск от спонтанен аборт. След това разгледахме връзката поотделно при жени с и без множество предишни спонтанни аборти (≥2). Таблица 3 показва, че асоциацията е до голяма степен сходна между тези две групи, като асоциацията е малко по-силна сред жените без множество предишни спонтанни аборти.
Таблица 3 Излагане на силни магнитни полета (МФ) по време на бременност и риск от спонтанен аборт, стратифицирано по брой предишни спонтанни аборти, МФ, измерено само в типични дни . Таблица 4 показва възможната връзка доза-отговор чрез изследване на връзката за всеки квартил, като се използва най-ниският квартил (2,5 mG) като референтна група. Докато всички по-високи квартили показват повишен риск от спонтанен аборт в сравнение с групата с най-ниска експозиция на MF, не се наблюдава връзка доза-отговор. Тези резултати са подобни на тези от предишно проучване 12 .
Таблица 4 Излагане на силни магнитни полета (МФ) по време на бременност и риск от спонтанен аборт - Оценка на реакцията на дозата, МФ, измерена само в типични дни . Наблюдаваната по-горе асоциация е последователна, независимо от източника на MF. Въпреки че не разполагахме с информация за точните източници, от които е генериран MF, въз основа на дневника на участниците, успяхме да проверим дали експозицията на MF е от някоя от следните категории местоположение: у дома, у дома в леглото, на работа, при транзитно преминаване или от други източници. Асоциацията се наблюдава последователно, независимо от местоположението. В допълнение към коригираните променливи, споменати по-горе, по-нататъшното приспособяване за гадене и повръщане, както и следните променливи не променят резултатите в таблици 2 - 4: доходи на майката, семейно състояние, гадене / повръщане при майката, употреба на алкохол, прием на кофеин, майчина треска, вагинално кървене, инфекция на пикочните пътища, носене на товари> 10 паунда, излагане на разтворители или обезмаслители, прием на витамини и джакузи / гореща вана / пара използване на стая / сауна по време на бременност.
Дискусия
След първоначални доклади, които предоставят доказателства за повишен риск от спонтанен аборт, свързан с висока експозиция на МФ по време на бременност 12 , 13 , настоящото финансирано от NIEHS проучване предоставя допълнителни доказателства, че излагането на високи нива на МФ по време на бременност е свързано с повишен риск от спонтанен аборт Тази констатация се подкрепя и от четири други проучвания, публикувани през последните 15 години, които изследват връзката между високата експозиция на МФ и риска от спонтанен аборт 8 , 9 , 10 , 11 , 19. Две от тези проучвания измерват ЕМП както вътре, така и в околните райони на пребиваването на участващите бременни жени и наблюдават по-висок риск от спонтанен аборт, свързан с по-високи нива на експозиция на ЕМП 8 , 9 . Две други проучвания изследват въздействието на ЕМП, излъчвани от клетъчни телефони и безжични мрежи, и установяват, че по-честата употреба на мобилен телефон и непосредствената близост до безжичните базови станции са свързани с повишен риск от спонтанен аборт 10 , 11 . Въпреки че нито едно от тези проучвания не е извършило никакви лични измервания на MF, за да улови действителната експозиция на MF от всички източници, както прави настоящото проучване, и четирите проучвания съобщават за повишен риск от спонтанен аборт, свързан с висока експозиция на MF.
Един от най-предизвикателните аспекти на оценката на въздействието на МФ върху здравето е способността да се измерва експозицията на МФ точно, както и в съответния етиологичен период. Проспективното измерване на експозицията на MF в етиологично значимия период е от съществено значение и за предпочитане пред ретроспективните измервания. Особено проблематично е да се установи експозицията на MF дълго след преминаване на съответния прозорец на експозиция. Макар и логистично предизвикателство, дизайн на проспективно проучване с устройство, което улавя действителните нива на MF от всички излъчващи източници в етиологично значим период, значително ще подобри точността на оценката на експозицията на MF при епидемиологични проучвания при човешка популация. В допълнение, както това проучване, така и предишно проучване 12доказано, дори и с перспективен дизайн, ако измерванията не са били проведени в обикновен ден, за да отразят истинската експозиция на MF по време на бременност, такъв дизайн на изследването все още не може да открие какъвто и да е риск за здравето на MF поради неправилна класификация на експозицията на MF (вж. таблица 2 ) Следователно, за да се осигури точна оценка на експозицията, MF измерванията трябва да се извършват проспективно по време на етиологично значим прозорец ида отразява типичните модели на експозиция на MF на участника. Определянето дали моделът на активност е типичен трябва да бъде проверен след приключване на измерването, тъй като планираните дейности могат да се променят по време на деня на измерване. Ясно е, че ако експозицията на MF се измерва субективно (напр. Интервю въз основа на отзоваването на участниците) или на базата на заместващи мерки (напр. Телени кодове, разстояние от електропроводи, матрица на работа, точково измерване у дома и т.н.), то за такива проучвания би било много трудно да се открие какъвто и да е здравен ефект в епидемиологичните проучвания поради груби неточности при измерване на действителните нива на експозиция на МФ. По дефиниция неточните MF мерки водят до погрешно класифициране на експозицията на MF, което обикновено води до нищожни констатации. За жалост, по-голямата част от епидемиологичните проучвания на ефектите върху здравето на MF в литературата досега се основават на субективни и ненадеждни измервания на MF. Поради това не е изненадващо, че много от миналите проучвания не успяха да открият MF здравни ефекти. В допълнение, фокусът върху изучаването на ефектите на МФ върху рака изостри проблема, тъй като развитието на рак обикновено има дълъг период на латентност между експозицията и резултата, който може да обхване няколко десетилетия. Това направи почти невъзможно точното измерване на експозицията на МФ в етиологично значимия период (десетилетия преди диагностицирането на рак). Тези „нулеви констатации“ са оставили фалшиво впечатление за „безопасността“ на експозицията на MF. фокусът върху изучаването на ефектите на MF върху рака изостри проблема, тъй като развитието на рак обикновено има дълъг латентен период между експозицията и резултата, който може да обхване няколко десетилетия. Това направи почти невъзможно точното измерване на експозицията на МФ в етиологично значимия период (десетилетия преди диагностицирането на рак). Тези „нулеви констатации“ са оставили фалшиво впечатление за „безопасността“ на експозицията на MF. фокусът върху изучаването на ефектите на MF върху рака изостри проблема, тъй като развитието на рак обикновено има дълъг период на латентност между експозицията и резултата, който може да обхване няколко десетилетия. Това направи почти невъзможно точното измерване на експозицията на МФ в етиологично значимия период (десетилетия преди диагностицирането на рак). Тези „нулеви констатации“ са оставили фалшиво впечатление за „безопасността“ на експозицията на MF.
Силата на настоящото проучване е, че в допълнение към използването на обективно измервателно устройство (измервател EMDEX Lite), ние изследвахме резултат (спонтанен аборт) с кратък период на латентност (дни или седмици, а не години или десетилетия, както в случая на ракови заболявания) или автоимунни заболявания). По този начин успяхме да измерим експозицията на MF проспективно в съответния период от време (по време на бременност). Освен това в края на деня на измерване установихме дали моделите на активност през този ден отразяват типичен ден, което ни позволи да идентифицираме участниците с измервания на експозиция на МФ, които по-точно отразяват експозицията на МФ по време на тяхната бременност.
В това проучване открихме почти трикратно повишен риск от спонтанен аборт, ако бременна жена е била изложена на по-високи нива на МФ в сравнение с жени с по-ниска експозиция на МФ. Асоциацията беше независима от каквито и да било източници или местоположения на експозиция на MF, като по този начин премахна опасението, че други фактори, свързани с източниците на експозиция, могат да отчетат наблюдаваните асоциации. Докато се предполагаше, че гаденето и повръщането представляват потенциални смущения, корекцията както за гадене, така и за повръщане не променя резултатите в това проучване или в предишно проучване 20. Въпреки че не наблюдавахме съотношение доза-отговор за експозиция на MF над 2,5 mG, това може да се дължи на прагов ефект от експозицията на MF, при който нивата на MF при или над 2,5 mG могат да доведат до смърт на плода, като по този начин се изследват допълнителни по-високи нива на MF експозицията не са в състояние да дадат допълнителен риск.
Като се има предвид повсеместният характер на излагане на това нейонизиращо лъчение, малък повишен риск, дължащ се на излагане на МФ, може да доведе до неприемливи последици за здравето на бременните жени. Въпреки че броят на епидемиологичните проучвания, изследващи неблагоприятното въздействие на експозицията на MF при хората, остава ограничен, резултатите от това проучване трябва да насочат вниманието към тази потенциално важна опасност за околната среда за бременни жени, поне в контекста на риска от спонтанен аборт, и да стимулират така необходимите допълнителни изследвания.
Препратки
- 1.
Wyde, М. и сътр . Доклад за частични констатации от Националната токсикологична програма Изследвания за карциногенеза на радиочестотна радиация на клетъчен телефон при Hsd: Sprague Dawley® SD плъхове ( Цялостно излагане на тялото ), http://biorxiv.org/content/early/2016/06/23/055699 ( 2016).
- 2.
Национална програма по токсикология. Медиен телебрифинг : Изследване за радиочестотна радиация на клетъчния телефон на NTP : Частично публикуване на констатациите , http://www.niehs.nih.gov/news/newsroom/releases/2016/may27/ (2016).
- 3.
Baan, R. et al . Канцерогенност на радиочестотните електромагнитни полета. The Lancet. Онкология 12 , 624–626 (2011).
- 4.
Международна агенция за изследване на рака Работна група за оценка на канцерогенните рискове за хората. Нейонизиращо лъчение , Част 2: Радиочестотни електромагнитни полета . Кн. 102 (Световна здравна организация, 2013).
- 5.
Световна здравна организация. 2007 Програма за изследване на СЗО за полета с изключително ниска честота. (Световната здравна организация, Женева, Швейцария, 2007 г.).
- 6.
Lindbohm, ML et al . Магнитни полета на видео дисплей терминали и спонтанен аборт. Am.J.Epidemiol. 136 , 1041–1051 (1992).
- 7.
Juutilainen, J., Matilainen, P., Saarikoski, S., Laara, E. & Suonio, S. Ранна загуба на бременност и излагане на 50-Hz магнитни полета. Биоелектромагнетика 14 , 229–236 (1993).
- 8.
Wang, Q. et al . Излагане на жилища на 50 Hz магнитни полета и връзката с риска от спонтанен аборт: 2-годишно проспективно кохортно проучване. PLoS One 8 , e82113 (2013).
- 9.
Shamsi, MF, Ziaei, S., Firoozabadi, M. & Kazemnejad, A. Излагане на изключително нискочестотни електромагнитни полета по време на бременност и риск от спонтанен аборт: Проучване за контрол на случая. J Res Health Sci 13 , 131–134 (2013).
- 10.
Zhou, LY et al . Епидемиологично изследване на рисковите фактори на бременните жени с ранен спонтанен аборт в Пекин. Chin J Integr Med . https://doi.org/10.1007/s11655-015-2144-z (2015).
- 11.
Mahmoudabadi, FS, Ziaei, S., Firoozabadi, M. & Kazemnejad, A. Използване на мобилен телефон по време на бременност и риск от спонтанен аборт. J Environment Health Sci Eng 13 , 34, https://doi.org/10.1186/s40201-015-0193-z (2015).
- 12.
Li, DK и сътр . Популационно проспективно кохортно проучване на лична експозиция на магнитни полета по време на бременност и риск от спонтанен аборт. Епидемиология 13 , 9–20 (2002).
- 13.
Lee, GM, Neutra, RR, Hristova, L., Yost, M. & Hiatt, RA Вложено изследване за контрол на случаи на жилищни и лични мерки на магнитно поле и спонтанни аборти. Епидемиология 13 , 21–31 (2002).
- 14.
Su, XJ et al . Корелация между излагането на магнитни полета и ембрионалното развитие през първия триместър. PLoS One. 9 , e101050 (2014).
- 15.
Гордън, НП Сравнение на социодемографските и здравните характеристики на членството на Kaiser Permanente в Северна Калифорния, получено от два източника на данни: Проучването за здравето на членовете от 2008 г. и Проучването за интервюто за здравето в Калифорния от 2007 г. (Kaiser Permanente, Изследователски отдел, Оукланд, Калифорния, 2012) .
- 16.
Гордън, НП Прилика на членството за възрастни Kaiser Permanente в Северна Калифорния със застрахованото и общото население в Северна Калифорния: Статистика от анкетата за здравно интервю в Калифорния 2011–12. (Отдел за изследвания на Kaiser Permanente, Оукланд, Калифорния, 2015 г.).
- 17.
Li, DK, Chen, H. & Odouli, R. Излагане на майката на магнитни полета по време на бременност във връзка с риска от астма при потомството. Арх . Педиатър . Юношеска . Med . (2011).
- 18.
Li, DK, Ferber, JR, Odouli, R. & Quesenberry, CP Jr. Проспективно проучване на вътреутробната експозиция на магнитни полета и риска от детско затлъстяване. Sci.Rep. 2 , 540 (2012).
- 19.
Shah, SG & Farrow, A. Систематичен преглед на литературата на неблагоприятните репродуктивни резултати, свързани с професионалното излагане на физиотерапевтите на нейонизиращо лъчение. J Заемане . Здраве (2014).
- 20.
Li, DK & Neutra, RR Магнитни полета и спонтанен аборт. Епидемиология 13 , 237–238 (2002).
Благодарности
Това проучване беше подкрепено със средства, предоставени от Националния институт по екологични здравни науки ( NIEHS ), безвъзмездна помощ № 5 R01 ES12226–5. Някои от резултатите бяха представени на годишни конференции на Международното общество по екологична епидемиология и общество за биоелектромагнетика 2015 и 2016 г.
Информация за автора
Принадлежности
Вноски
De-Kun Li замисля концепцията, проектира проучването, получава финансиране, ръководи събирането на данни и анализите и отговаря за интерпретацията на резултатите, както и за изготвянето и финализирането на ръкописа. Жанет Фербер и Хонг Чен отговаряха за управлението на данните. Хонг Чен участва в анализа на данните и интерпретацията на резултатите. Роксана Одули участва в ръководството на изследването и подготовката на ръкописа. Чарлз Куесенбъри участва в интерпретацията на резултатите и подготовката на ръкописа. De-Kun Li е гарантът на този документ, който пое цялата отговорност за провеждането на проучването, имаше достъп до данните и контролира решението за публикуване.
Автора за кореспонденция
Етични декларации
Конкуриращи се интереси
Авторите заявяват, че нямат конкуриращи се интереси.
Допълнителна информация
Бележка на издателя: Springer Nature остава неутрален по отношение на юрисдикционни претенции в публикувани карти и институционални принадлежности.
- 1.
Ефекти от развитието след поведението и поведението при вътреутробно излагане на плъхове на радиочестотни вълни, излъчвани от конвенционални WiFi устройства. Othman, H., et al. Токсикология и фармакология на околната среда 52: 239-247 (2017).
тотни вълни, излъчвани от конвенционални WiFi устройства
Принадлежности- PMID: 28458069
- DOI: 10.1016 / j.etap.2017.04.016
Резюме
Настоящата работа изследва ефектите от пренаталното излагане на радиочестотни вълни на конвенционални WiFi устройства върху постнаталното развитие и поведението на потомството на плъхове. Десет бременни плъхове с албинос от Wistar са разпределени на случаен принцип в две групи (n = 5). Експерименталната група беше изложена на 2,45 GHz WiFi сигнал в продължение на 2 часа на ден през целия период на бременността. Контролните жени са били подложени на същите условия като третираната група, без да се прилага WiFi лъчение. След раждането, потомството е тествано за физическо и невроразвитие през своите 17 постнатални дни (PND), след това за тревожност (PND 28) и моторност (PND 40-43), както и за церебрална реакция на оксидативен стрес и активност на холинестераза в мозъка и серум (PND 28 и 43). Нашите основни резултати показват, че вътреутробната експозиция на WiFi е нарушила невроразвитието на потомството през първите седемнадесет постнатални дни, без да променя емоционалното и двигателното поведение в зряла възраст. Освен това, пренаталната експозиция на WiFi предизвиква дисбаланс на мозъчния оксидативен стрес (повишаване на нивото на малондиалдехид (MDA) и водороден пероксид (H2 O 2 ) нива и намаляване на активността на каталазата (CAT) и супероксиддисмутазата (SOD) на 28, но не на 43 дни, също експозицията засяга активността на ацетилколинестеразата както на церебрално, така и на серично ниво. По този начин, настоящото проучване разкрива, че излагането на майката на WiFi радиочестоти води до различни неблагоприятни неврологични ефекти при потомството, като влияе върху невроразвитието, равновесието на мозъчния стрес и холинестеразната активност.
Ключови думи: поведение; Потомство; Оксидативна реакция на стрес; Постнатално развитие; Радио честота.
Copyright © 2017 Elsevier BV Всички права запазени.
Подобни статии
- Ефекти от пренаталното излагане на WIFI сигнал (2,45 GHz) върху постнаталното развитие и поведение при плъхове: Влияние на сдържаността на майката.Behav Brain Res. 2017 май 30; 326: 291-302. doi: 10.1016 / j.bbr.2017.03.011. Epub 2017 10 март.PMID: 28288806
- Ефекти на повтарящ се стрес и експозиция на WiFi сигнал върху поведението и оксидативен стрес при плъхове.Metab Brain Dis. 2017 октомври; 32 (5): 1459-1469. doi: 10.1007 / s11011-017-0016-2. Epub 2017 27 април.PMID: 28451780
- Пренаталното приложение на липополизахариди при майките води до специфичен за възрастта и региона оксидативен стрес в ранния етап на развитие при потомството.Неврология. 2016 г. 24 март; 318: 84-93. doi: 10.1016 / j.neuroscience.2016.01.002. Epub 2016 8 януари.PMID: 26774051
- Оксидативен стрес и инхибиране на холинестеразата в плазмата, еритроцитите и мозъка на малките плъхове след лактационно излагане на малатион.Environment Toxicol Pharmacol. 2012 ноември; 34 (3): 753-60. doi: 10.1016 / j.etap.2012.09.012. Epub 2012 28 септември.PMID: 23122842
- Добавянето на витамин А при плъхове по време на бременност и кърмене предизвиква поведенчески промени и оксидативен стрес върху стриатума и хипокампуса на майките и техните потомци.Brain Res. 2011 г. 19 януари; 1369: 60-73. doi: 10.1016 / j.brainres.2010.11.042. Epub 2010 17 ноември.PMID: 21092734
Цитирано от 3 статии
- Ранното излагане на импулсни LTE радиочестотни полета причинява трайни промени в активността и поведението на мишки C57BL / 6 J.Биоелектромагнетика. 2019 октомври; 40 (7): 498-511. doi: 10.1002 / bem.22217. Epub 2019 15 септември.PMID: 31522469 Безплатна статия от PMC.
- Ефект на микровълнова Wi-Fi радиация при честота 2,4 GHz върху епилептичното поведение на плъховете.J Biomed Phys Eng. 2018 юни 1; 8 (2): 185-192. eCollection 2018 юниPMID: 29951445 Безплатна статия от PMC.
- Последен напредък в ефектите на микровълновото лъчение върху мозъка.Mil Med Res. 2017 г., 21 септември; 4 (1): 29. doi: 10.1186 / s40779-017-0139-0.PMID: 29502514 Безплатна статия от PMC. Преглед.
Условия на MeSH
Вещества
LinkOut - повече ресурси
Източници на пълен текст
Разни
Трайни хепатотоксични ефекти от пренаталната експозиция на мобилен телефон. Yilmaz, A., et al. Вестник по майчина-фетална и неонатална медицина 30 (11): 1355-1359 (2017).
Трайни хепатотоксични ефекти на пренаталната експозиция на мобилен телефон
Принадлежности- PMID: 27427155
- DOI: 10.1080 / 14767058.2016.1214124
Резюме
Цел: В това проучване черният дроб на плъхове, родени от майки, изложени на електромагнитно поле (ЕМП), са изследвани 60 дни след раждането за биохимични и хистопатологични промени.
Методи: Бременните плъхове са били изложени на радиация (900 MHz EMF, 24 часа / ден в продължение на 20 дни) с помощта на цифров генератор на сигнали чрез поставяне на устройството централно под клетката, която е образувала изследваната група (EMF), докато нетретираните плъхове, които отговарят контроли. Черният дроб и кръвта са получени от носилки (седем мъже и седем жени) от двете групи 60 дни след раждането, които са били използвани за биохимични и хистопатологични анализи.
Резултати: Налице е значително увеличение на нивата на малондиалдехид (MDA) (p <0,05), което е придружено от значителен спад в глутатиона (GSH) (p <0,01) в черния дроб. Серумните нива на аланин аминотрансфераза (ALT) и аспартат аминотрансфераза (AST) са значително повишени (р <0,05). Хистопатологично чернодробните участъци от групата с ЕМП показват интензивна дегенерация в хепатоцити с цитоплазмени еозинофилни структури, пикнотични ядра и фиброза.
Заключение: Демонстрираме, че вътрематочното вредно въздействие на ЕМП върху черния дроб на плъховете продължава и до зряла възраст.
Ключови думи: Излъчване на мобилен телефон; електромагнитно поле; чернодробна токсичност; пренатална експозиция.
Подобни статии
- Пренатални ефекти от 1800 MHz електромагнитно поле върху черния дроб на плъхове.Клетки тъканни органи. 2019; 207 (3-4): 187-196. doi: 10.1159 / 000504506. Epub 2019 4 дек.PMID: 31801129
- Излагането на майката на непрекъснато 900-MHz електромагнитно поле провокира невронална загуба и патологични промени в малкия мозък на 32-дневно женско потомство на плъхове.J Chem Neuroanat. 2016 г. септември; 75 (Pt B): 105-10. doi: 10.1016 / j.jchemneu.2015.09.002. Epub 2015 21 септември.PMID: 26391347
- Ефект на излагане и отнемане на 900-MHz електромагнитни вълни върху мозъчния, бъбречния и чернодробния оксидативен стрес и някои биохимични параметри при мъжки плъхове.Electromagn Biol Med. 2015; 34 (4): 279-84. doi: 10.3109 / 15368378.2014.906446. Epub 2014 8 април.PMID: 24712749
- Вредни ефекти от дългосрочно, непрекъснато 900-MHz електромагнитно поле през юношеството върху морфологията на хипокампуса, биохимията и броя на пирамидалните неврони при мъжки плъхове на Sprague Dawley на 60 дни.J Chem Neuroanat. 2016 ноември; 77: 169-175. doi: 10.1016 / j.jchemneu.2016.07.004. Epub 2016 16 юли.PMID: 27430379
- Ефектите от пренаталното дълготрайно излагане на 900-MHz електромагнитно поле върху 21-дневния новороден черен дроб на мъжки плъх.Turk J Med Sci. 2015; 45 (2): 291-7. doi: 10.3906 / sag-1404-168.PMID: 26084117
Цитирано от 3 статии
- Ефектите от боровинки върху индуцирана от исхемия реперфузия на миокардно увреждане при плъхове.Турчин Гогус Калп Дамар Черасиси Дерг. 2020 г., 23 януари; 28 (1): 63-69. doi: 10.5606 / tgkdc.dergisi.2020.18389. eCollection 2020 януариPMID: 32175144 Безплатна статия от PMC.
- Радиозащитният ефект на N-ацетилцистеин срещу индуцирано от рентгеново лъчение бъбречно увреждане при плъхове.Environment Sci Pollut Res Int. 2019 октомври; 26 (28): 29085-29094. doi: 10.1007 / s11356-019-06110-0. Epub 2019 7 август.PMID: 31392607
- Уортберили предпазва бъбреците срещу индуцираната от цисплатин нефротоксичност: експериментално проучване.Ren Fail. 2018 ноември; 40 (1): 466-474. doi: 10.1080 / 0886022X.2018.1500292.PMID: 30130136 Безплатна статия от PMC.
Условия на MeSH
Вещества
LinkOut - повече ресурси
Източници на пълен текст
Медицински
Многобройни методи за оценка на пренаталното излагане на радиочестотно излъчване от телекомуникациите в изследването на майките и здравето на околната среда на децата (MOCEH). Choi, Ha, et al. Международен вестник по трудова медицина и здраве на околната среда 29 (6): 959-972 (2016).Int J Occup Med Environment Health
doi: 10.13075 / ijomeh.1896.00803. Epub 2016 3 ноември.. 2016 г. 18 ноември; 29 (6): 959-972.Многобройни методи за оценка на пренаталното излагане на радиочестотно излъчване от телекомуникациите в изследването на майките и здравето на околната среда (MOCEH)
Принадлежности- PMID: 27869246
- DOI: 10.13075 / ijomeh.1896.00803
Резюме
Цели: Да се оцени пренаталното излагане на радиочестотно излъчване (RFR) от далекосъобщенията с помощта на въпросник за мобилен телефон, дневници на данни на оператора за използване на мобилни телефони и измервателен уред за персонална експозиция (PEM).
Материали и методи: Проучването включва 1228 двойки майки и бебета от проучването на майките и здравето на околната среда (MOCEH) - многоцентрово проспективно кохортно проучване, продължаващо от 2006 г., в което участниците са записани на ≤ 20 седмици от бременността, с проследяване на раждането на дете и растеж, за да се оцени връзката между пренаталното излагане на околната среда и здравето на децата. Въпросникът включваше средната честота на разговорите на ден и средното време за разговори на ден. EME Spy 100 PEM беше използван за измерване на RFR сред 269 бременни жени от ноември 2007 г. до август 2010 г. Данните от операторите бяха получени от 21 участници. Тестът за корелация на Спиърман е извършен за оценка на коефициента на корелация и 95% доверителни интервали между информацията за използване на мобилния телефон от въпросника,
Резултати: Регистрационните данни на операторите и информацията от самоотчетения въпросник показват значително високи корелации в средната честота на разговорите на ден (ρ = 0,6, p = 0,004) и средното време на повикване на ден (ρ = 0,5, p = 0,02) . Корелацията между информацията за употребата на мобилен телефон в самоотчетения въпросник и индекса на експозиция, регистриран от PEM, беше лоша. Но корелацията между информацията от регистрационните данни на операторите и индекса на експозиция за предаване на мобилна комуникация беше значително висока: коефициентът на корелация (р-стойност) беше 0,44 (0,07) за честотата на повикване на ден и беше 0,49 (0,04) за повикването време на ден.
Заключения: Информацията за въпросника за използването на мобилния телефон показва умерено до високо качество. Използването на множество методи за оценка на експозицията може да е по-добро от използването само на един метод. Int J Occup Med Environ Health 2016; 29 (6): 959-972.
Ключови думи: MOCEH; мобилен телефон; измервателен уред за лична експозиция; пренатална експозиция; радиочестотно излъчване; валидиране.
Това произведение се предлага в модел с отворен достъп и се лицензира под лиценз CC BY-NC 3.0 PL.
Подобни статии
- Невроразработка за първите три години след пренатална употреба на мобилен телефон, радиочестотно излъчване и излагане на олово.Environment Res. 2017 юли; 156: 810-817. doi: 10.1016 / j.envres.2017.04.029. Epub 2017 13 май.PMID: 28511138
- Пренатален бисфенол А и резултати от раждането: проучване MOCEH (Майки и здраве на околната среда на децата).Int J Hyg Environment Health. 2014 март; 217 (2-3): 328-34. doi: 10.1016 / j.ijheh.2013.07.005. Epub 2013 11 юли.PMID: 23911140
- Валидиране на оценката на експозицията и оценка на методите за набиране за проспективно кохортно проучване на потребители на мобилни телефони (COSMOS) във Финландия: пилотно проучване.Environment Health. 2011 г. 8 март; 10:14. doi: 10.1186 / 1476-069X-10-14.PMID: 21385407 Безплатна статия от PMC.
- Ефект на радиочестотната радиация върху репродуктивното здраве.Indian J Med Res. 2018 декември; 148 (Suppl): S92-S99. doi: 10.4103 / ijmr.IJMR_1056_18.PMID: 30964085 Безплатна статия от PMC. Преглед.
- Систематичен преглед на здравните ефекти от излагането на радиочестотни електромагнитни полета от базовите станции на мобилните телефони.Bull World Health Organ. 2010 декември 1; 88 (12): 887-896F. doi: 10.2471 / BLT.09.071852. Epub 2010 5 октомври.PMID: 21124713 Безплатна статия от PMC. Преглед.
Цитирано от 1 статия
- Асоциации на използването на мобилен телефон на майката по време на бременност с продължителност на бременността и растеж на плода в 4 кохорти при раждане.Am J Epidemiol. 2019 юли 1; 188 (7): 1270-1280. doi: 10.1093 / aje / kwz092.PMID: 30995291 Безплатна статия от PMC.
Видове публикации
Условия на MeSH
LinkOut - повече ресурси
Източници на пълен текст
Изследователски материали
Използването на сигнално-трансдукционни и метаболитни пътища за прогнозиране на цели за човешка болест от електрически и магнитни полета, като се използват данни in vitro в човешките клетъчни линии. Parham, Portier, et al. Граници в общественото здраве (2016).
Преглед на електромагнитните полета (ЕМП) и репродуктивната система. Asghari, Khaki, et al. Електронен лекар 8 (7): 2655-2662 (2016).
Преглед на електромагнитните полета (ЕМП) и репродуктивната система
1. Въведение
Хората в съвременния свят често са изложени на електромагнитни полета (ЕМП). Излагането на хора на ЕМП идва от много източници и ситуациите са различни в ежедневието на хората. ЕМП произлизат от електропроводи, компютърни устройства, телевизори, радиостанции и телефони. Има много фактори, които влияят върху степента, до която хората могат да бъдат засегнати от ЕМП. Например телесното тегло, индексът на телесна маса, костната плътност и нивата на вода и електролити могат да променят проводимостта и биологичната реактивност на ЕМП ( 1 , 2). Следователно ефектите от това замърсяване на околната среда могат да зависят от пола, тъканната плътност на тялото, периода на живот и нивата на излагане на ЕМП. Започвайки през 1960 г., когато за първи път са изследвани биологичните опасности, причинени от ЕМП, човешкото здраве стана важен фокус на работното място и у дома ( 3 ). Въпреки че биологичните ефекти на ЕМП все още са противоречиви, като цяло отрицателните ефекти не трябва да се пренебрегват. В момента хората са изложени на различни видове ЕМП, които са нейонови лъчения, които не могат да отделят електрони. Те са енергия под формата на трептящи електрически и магнитни полета, които се трансформират от една точка в друга. Много форми на физическа енергия, като рентгенови лъчи, UV светлина и слънчева светлина, произвеждат ЕМП ( 4). Има няколко препратки, които класифицират ЕМП, но като цяло могат да се считат, че се състоят от четири различни типа. Първият тип електромагнитни полета се отнася до електромагнитни полета с изключително ниска честота (електромагнитни полета), които са електромагнитни полета, които са под 300 HZ и се произвеждат от военно оборудване и железопътни линии. Вторият тип, известен като EMF с междинна честота (IF), има честоти в диапазона от 300 Hz до 10 MHZ и се произвежда от индустриални кабели и електрическо оборудване в домовете, като телевизори и компютърни монитори. Третият тип е EMF с високочестотни честоти (HF), които имат честоти в диапазона от 10 MHz до 3000 GHz и се произвеждат от мобилни телефони и радиоразпръскване. Радиочестотите (RF) също са част от тази категория, която има честоти до 100 MHz ( 4). Има и статични ЕМП, които се получават чрез ЯМР и геомагнетизъм и са посочени с нулева честота ( 3 ). През 1979 г. Wertheimer и Looper показват, че съществува пряка връзка между ЕМП и повишената честота на левкемия при бебета ( 5 ). Ако биологичната система на тялото е изложена на ЕМП, които произвеждат електрически токове и полета, които всъщност се справят с тока и напрежението, нормалният физиологичен баланс се нарушава. Ако плътността на електрическия ток се увеличи до прага на стимулация, може да се получи мембранна деполяризация на нервите и мускулите. Електрическите и магнитните полета на нива на околната среда могат да удължат живота на свободните радикали и да доведат до увреждане на дезоксирибонуклеиновата киселина (ДНК) на хората ( 6). Някои епидемиологични проучвания са направени при различни популации, но повечето са направени върху лабораторни животни и клетъчни линии ( 4 ). Биологичните ефекти на ЕМП обикновено могат да бъдат разделени на топлинни и нетермични ефекти ( 7 ). Термичните ефекти се определят като топлината, генерирана от ЕМП в определена област. Нетермичните ефекти зависят от абсорбцията на енергия и промените в поведението на тъканите, без да произвеждат топлина. ЕМП имат висока проникваща сила и те са способни да движат заредени частици, като електроните и йоните на големи макромолекули и полимери ( 7 ). Така че ЕМП могат да имат опустошителни ефекти върху тъканите с високи концентрации на електрони и йони. ЕМП, които причиняват промени в поведението на клетките ( 8) и тъканите променят функцията на сърдечно-съдовата система ( 9 ) и костния мозък ( 7 ). Електромагнитните полета могат да имат няколко различни ефекти върху клетъчните компоненти ( 10 ), включително нарушения на клетъчната пролиферация и диференциация ( 10 ), увредена ДНК в клетките, хромозомни аномалии ( 11 ), кръвни нарушения ( 9 ), вродени дефекти ( 12 ) и различни мутации, включително тези, свързани с дългосрочно излагане на ЕМП. Под въздействието на тези полета балансът на ЦНС и хормоналната и дихателната системи стават слаби, което води до намалена активност на споменатите органи ( 13 , 14). Изследванията върху ефектите на ЕМП върху ендокринната система се фокусират най-вече върху мелатонина и производния триптофан, произведен от епифизната жлеза ( 15 ). Повечето от вредните ефекти на ЕМП действат чрез процеса на протеинов синтез ( 16 , 17 ). В това отношение ензимите, поради комбинацията от аминокиселини, са засегнати и тяхната каталитична активност е намалена ( 4 ). Проучванията относно ефектите на цитотоксичността и генотоксичността на ЕМП се фокусират най-вече върху фибробластите, меланоцитите, лимфоцитите, моноцитите и мускулните клетки при хора и върху гранолозните клетки на плъхове ( 18). Декласифициран доклад на агенцията за отбранително разузнаване от 1976 г. показва, че военният персонал, изложен на нетермична микровълнова радиация, изпитва „главоболие, умора, световъртеж, раздразнителност, безсъние, депресия, безпокойство, забрава и липса на концентрация ( 19 ). Проучване от A2015 показа, че 2,4 GHz WiFi може да бъде един от основните рискови фактори за мозъчни тумори и други невродегенеративни заболявания ( 20 ). Друга статия от 2015 г. показва, че поляризираната ЕМП (създадена от човека) е много по-активна биологично от неполяризираната ЕМП ( 21 ). Друга статия показва, че зайците изпитват сърдечна аритмия и повишено кръвно налягане, когато са изложени на Wi-Fi от 2,4 GHz ( 22). Дългосрочно проучване, проведено от Lennart Hardell, шведски учен, върху глиома и акустични невромни мозъчни тумори показа, че RF е канцерогенен. Ученият призова RF да бъде етикетиран като IARC клас 1 канцероген и препоръча спешно преразглеждане на насоките за безопасност ( 23 ). Проучване от 2011 г. на Нора Волкоу показа, че лъчението от мобилни телефони в зоните до антената повишава метаболизма на глюкозата в мозъка. Повишеният метаболизъм на глюкозата е свързан с рак. Изследването показа, че биологичните промени се случват на нива, по-ниски от настоящите насоки на FCC ( 24 ).
2. Дискусия
2.1. Мъжка полова система и ЕМП
Разрушителните ефекти на ЕМП върху двата пола са изследвани при различни експозиции ( 25 ). Тъй като репродуктивната система се контролира от нервната и ендокринната системи, замърсителите на околната среда, като EMF, могат да повлияят на двете системи, споменати по-горе, така че гениталната система също участва ( 4 ). Проучванията върху връзката между женските плъхове и неврордокринните промени показват, че невроендокринните нарушения са основната причина за проблеми с плодовитостта ( 26 ). Проучване от 2015 г. показа, че излагането на Wi-Fi от 2,4 GHz намалява функцията на сперматозоидите. Изследователите заключават, че "трябва да има сериозна загриженост във връзка с излагането на безжични мрежи, които съществуват в околностите на нашите живи места." ( 27). EMFs индуцират клетъчна смърт в тестикуларната зародишна клетка при мишки ( 28 ). През 2004 г. отрицателните ефекти на ЕМП включват промяна в серумните нива на тестостерон ( 29 - 31 ). Изследване на тестикуларната тъкан показа, че броят на клетките в цикъла на сперматогенеза е намалял значително ( 32 ). Доклади от изследвания на епидидима, семенните везикули и простатната жлеза при пренатални плъхове, изложени на ЕМП, показват, че тези животни на възраст от четири месеца нямат промени в нивата на половите хормони или броя на сперматозоидите ( 4 ). Множествен достъп с кодово разделяне (CDMA) на EMF с честота 84 MHz не повлиява значително телесното тегло, тестисите, епидидима, броя на сперматозоидите или зародишната клетка на апоптоза при възрастни плъхове ( 33). Също така, възрастни плъхове, които са били изложени на 900-MHz EMFs в продължение на 30 минути пет дни в седмицата, не са имали значителни промени в теглото на тестисите си или относителния процент от общия контекст на тестикуларната интерстициална тъкан, но са имали намалени серумни нива на тестостерон без промени в нивата на техния лутеинизиращ хормон (LH) и фоликулостимулиращ хормон (FSH) ( 34 ). При плъхове, изложени на 1,95 GHz EMF в продължение на пет часа на ден в продължение на пет седмици, не се наблюдават промени в тъканта и параметрите на репродуктивната система, но броят на техните сперматозоиди се увеличава независимо от аномалии ( 4 ). При високи честоти, особено 2,45 GHz, EMF намаляват броя на лейдиговите клетки и увеличават апоптозата в семенните тубули при плъхове ( 35). Честотата на RF от 2,45 MHz води до отказ в ДНК клетките на тестисите ( 36 ). Освен това излагането на RF (900 MHz – 1,7 GHz) причинява разкъсвания на ДНК в ембрионални стволови клетки и епидидимни сперматозоиди на мишки ( 34 ). 60 Hz и 0,5 T EMFs причиняват апоптоза на сперматогония и предизвикват увреждане на ДНК, но няма значителна промяна в процента на живите клетки ( 37 ). РЧ лъчението може да намали оплождането на сперматозоидите при мъжете ( 38 ). Обратно, SLF-EMF не оказват влияние върху жизнеспособността и морфологията на сперматозоидите от нерези, но вътреклетъчните нива на калций са намалени в сравнение с контролната група и има намалена подвижност и оплождане ( 4). При зайци 50 Hz SLF-EMF могат да причинят промени в подвижността на сперматозоидите и да намалят тяхната жизнеспособност ( 39 ). Този процес се осъществява чрез намеса във вътреклетъчната калциева хомеостаза, което води до увреждане на функцията на сперматозоидите. Изследователите показват, че функцията на сперматозоидите е намалена от EMF (интензитет 1 mT) in vivo ( 40 ). Изследване на мъжки плъхове показва, че честота от 50 Hz за 8 h / ден в продължение на осем месеца причинява хистологични вариации на тестисите, които включват загуба на тегло и намаляване на средния диаметър на семенните тубули ( 4). В едно проучване със светлинен микроскоп се наблюдава, че при 50-Hz EMFs сперматогониалните клетки се отделят от зародишния епител. Това проучване показа, че броят на зрелите сперматозоиди в семенните тубули е намален и някои от клетките на сертоли са сплескани ( 41 ). Всички доклади споменават ефектите на ЕМП върху ултраструктурната тъкан на мъжката репродуктивна система при модели на хора и животни. Но за допълнително изясняване трябва да се проведат изследвания на сигналните пътища и молекулярната структура на клетките.
2.2. Женска полова система и ЕМП
Женската репродуктивна система е съвкупност от различни тъкани, които са свързани помежду си. Така че, ако се обсъждат ефектите на EMF върху женското безплодие, ефектите от този фактор на околната среда трябва да бъдат оценени върху различните части на гениталната система. Женската полова система е съставена от матката, яйчниците, фалопиевите тръби, освободените ооцисти и зародишните и соматичните клетки в тяхната тъкан. Според много изследователи невроендокринните промени, причинени от ЕМП, са ключов фактор за промяна на хормоналната функция и причиняват симптоми на безплодие при жените ( 26 ). Редица други изследователи са се фокусирали върху естрозния цикъл ( 42). Една от разликите между сексуалните цикли на жените и женските гризачи е, че женските цикли са завършени и връхът на естроген-прогестрон е отделен. При женските гризачи пикът на тези два хормона е едновременно. Така че може би пикът на естроген-прогестрон трябва да се има предвид за бъдещо разследване. Освен това много изследвания са фокусирани върху вредното въздействие на ЕМП върху гранулозните клетки на ооцитите. Апоптозата на тези клетки също е друг въпрос в много статии ( 43 ). По-долу са представени няколко доклада по този въпрос. Спонтанният аборт и аномалии на плода са два взаимосвързани въпроса, които привлякоха вниманието на много изследователи ( 44). Клиничните проучвания върху бременни жени, изложени на видео дисплей терминал (VDT), показват значително увеличение на спонтанните аборти ( 45 ). Съществува положителна връзка между професионалните монитори с бременна жена и съотношението на феталните аномалии ( 45 ). Епидемиологичните проучвания върху вродени дефекти и спонтанни аборти от заети бременни жени показват, че генерираните от ЕМП компютри могат да имат отрицателно въздействие върху репродуктивната система на хората ( 46 ). Въпреки че отрицателните ефекти върху развитието на плода са противоречиви, някои разрушителни фактори са докладвани в много животински модели ( 4). Мъжкото потомство на женски плъхове след излагането им на честота 60 Hz, интензитет 1 mT на SLF-EMF показва, че е имало намаляване на броя, височината и обема на семенните тубули, както и в броя и диаметъра на лейдиговите клетки ( 47 ). Въпреки това, серумните нива на тестостерон, гонадозоматичен индекс и сертоли клетки остават непроменени ( 47 ). За разлика от това, когато бременни плъхове са били изложени на SLF-EMF (честота 60 Hz, 500 μT интензивност) в продължение на 21 часа на ден, мощността на плодовитостта и сперматогенезата на мъжкото потомство не са били значително намалени в експерименталната група в сравнение с контролната група ( 48). Друго проучване за определяне на връзката между ЕМП и овулацията при плъхове показва, че вълните инхибират овулацията и намаляват броя на жълтите тела ( 49 ). Женските мишки, изложени на 20 KHz EMF от телевизори и компютри, са имали удължени естрални цикли, което е един от най-важните фактори за намаляване на цикъла на бременност при женските животни ( 50 ). Излагането на ЕМП предизвиква репродуктивен ефект и ефект на токсичност за развитието, който дегенерира ооцити при мишки ( 31 ). Култивираните фоликули в мишката след излагане с 33 Hz SLF-EMF причиняват поражение на растежа ( 4 ). За разлика от това, излагането на 55 Hz причинява продължителен растеж на фоликулите ( 51). Друго изследване показва, че излагането на 33-50 Hz в продължение на три дни предотвратява образуването на антрални фоликули in vitro ( 51 ). Възрастни женски плъхове, които са били изложени на 50 Hz честота на SLF, не са имали значителни промени в теглото на матката, яйчниците и нивата на прогестерон и естроген ( 52 ). След отстраняване на матката при женски плъхове и излагане на 1.439 MHz на EMF (TDMA) в продължение на 4 часа / ден, няма разлика в обема на матката и нивата на стероидни хормони ( 53 ). Когато женските плъхове бяха изложени на 900 MHz в продължение на 30 дни, апоптозата на ендометриума и оксидативният стрес се увеличиха ( 54). Отчетени са ефектите на EMF върху имплантацията и развитието на плода. По време на чифтосване, мишки, изложени на честота 50 Hz, интензитет 0,5 mT в продължение на 4 часа / ден в продължение на две седмици, са имали значително намаляване на броя на бластоцистите и увеличават фрагментацията на ДНК ( 55 ). Това проучване предполага, че излагането на EMF в периода на имплантиране може да има вредни ефекти върху развитието на ембрионите. Радиацията за 4 часа преди овулацията показа, че SLF-EMFs имат отрицателни ефекти върху ранното развитие на ембриона ( 4 ). Когато бременните мишки са били изложени на честота 50 Hz, интензитет 20 mT, степента на преживяемост, съотношението между половете и феталните малформации не са се променили значително, но височината и теглото на плодовете са се увеличили значително ( 56). Проучване върху ефектите на ЕМП върху женски плъхове, анализирано чрез трансмисионен електронен микроскоп (ТЕМ), показва увеличаване на броя на макрофагите и вакуолите на автофагията в някои гранулозни клетки и наличието на няколко липидни капки в лутеалните и тека клетките ( 56 ). Други изследователи са показали, че ЕМП причиняват повишени макрофаги в лутеа корпусите и нарастващите фоликули. Те вярвали, че EMF ускоряват процеса на апоптоза в яйчниците на плъхове. В допълнение, повечето изследователи смятат, че ЕМП чрез апоптоза причиняват разрушаването на кортикалната тъкан на яйчниците, луминалния епител, жлезистия епител и стромалните клетки в матката и фалопиевите тръби ( 57 ).
2.3. Определение на антиоксиданта
Обикновено аеробните организми имат различни защитни системи или антиоксидативни механизми за противодействие на щетите, причинени от реактивни кислородни видове (ROS) и техните продукти. Тези защитни механизми действат чрез предотвратяване на верижната реакция, инициирана от ROS или чрез прекъсване на верижната реакция след нейното иницииране. Антиоксидантите са съединения, които съдържат монохидрокси / полихидрокси фенол и са разделени на три групи, т.е. 1) първични антиоксиданти, които предотвратяват образуването на оксиданти; 2) вторични антиоксиданти, които възстановяват окислените молекули чрез диетични или последователни антиоксиданти; и 3) превантивни антиоксиданти, т.е. антиоксиданти, които свързват метални макромолекули (албумин, церулоплазмин и трансферин) и антиоксидантни ензими. Последователните антиоксиданти се състоят от разтворим в липиди разтворим витамин Е, убиквитон, β-каротин, водоразтворим GSH, аскорбат и урат. Антиоксидантите са от съществено значение за предотвратяване на сърдечни заболявания, рак и диабет (88). Клетките имат много антиоксиданти, които ги предпазват от увреждане. Предотвратяването на прекомерни ROS и възстановяването на клетъчни увреждания е от съществено значение за живота на клетката. В този преглед представяме два примера за естествени антиоксиданти, които са регистрирани в много проучвания, т.е. витамин Е и екстракт от копър.
2.4. Антиоксидантни свойства на витамин Е
Витамин Е е най-важният мастноразтворим антиоксидант, който се намира в мазнината на клетъчната мембрана и предпазва ненаситените фосфолипидни мембрани от свободните радикали и реакционното окисляване ( 54). Естествена форма на витамин Е е d-алфа токоферолът. Витамин Е елиминира свободните радикали, като ги превръща в безвредни метаболити. Най-важните дейности на витамин Е са, че той подобрява регенеративния цикъл, повишава еластичността и повишава толерантността на организма. Абсорбцията на витамин Е зависи от функцията на панкреаса и жлъчния секрет. Този витамин се пренася от липопротеините в плазмата и еритроцитите. Клетъчното усвояване на витамин Е се осъществява чрез два механизма, т.е. 1) процес, медииран от рецептори, при който LDL доставя витамина до клетките и 2) Процес, медииран от липопротеини, който разделя витамин Е от хиломикроните и VLDL. Витамин Е е един от фенолните антиоксиданти и дефицитът на този витамин може да доведе до неблагоприятни физически ефекти. Като цяло, дефицитът на витамин Е излага на риск нервно-мускулните, сърдечно-съдови и генитални системи. Витамин Е има значително влияние върху намаляването на абортите на плода. Положителното въздействие на витамин Е върху анатомичните и хистологичните аспекти на гениталната система е показано в многобройни проучвания (59 , 60 ).
2.5. Антиоксидантни свойства на копър
Копърът е от семейство магданоз (Apiaceae) и расте естествено в Европа и Средиземноморския регион; сега се култивира по целия свят. Обикновено два вида от това растение са годни за консумация, т.е. горчив и сладък копър. Той има приложения в традиционната медицина. В древен Китай и Египет се използва за лечение на ухапвания от змии, като храна и за медицински цели. Копърното масло има антиоксидантно, антимикробно, спазмолитично и стомашно-чревно стимулиращо действие ( 61 ). Алергичните реакции към него се появяват много рядко. Витамин С е най-активният компонент на копъра, съставлявайки 17% от него. Следователно, той има противовъзпалителни свойства и поради своите влакна предотвратява предаването на токсини в дебелото черво. В допълнение към влакната, копърът съдържа калциеви, магнезиеви, медни и фосфорни йони ( 61). Анетолът е най-основното съединение от копър, което намалява възпалението и предотвратява рака ( 62 ). Анетолът също предпазва черния дроб от увреждане. Изследователите приписват противовъзпалителните и противораковите свойства на копъра да отлагат или изключват вътреклетъчния сигнален път, наречен TNF ( 63 ). По този начин, копърът може да инхибира промените и елиминира възпалителните каскадни молекули, наречени NF-Kappa B ( 63 ). Копърът предизвиква естрозния процес при женски плъхове. Копърът драстично увеличава броя на антралните, графиовите и многослойните фоликули и подобрява процеса на фоликугенеза в яйчниците на мишки ( 61). Тъй като растежът на фоликулите се влияе от ендокринните хормони (FSH, LH, пролактин и паракринни фактори), естрогенните ефекти на копъра могат да увеличат броя на фоликулите, които растат ( 61 ). Друга активна съставка на копъра е диосгенинът, който е от съществено значение за синтеза на някои хормони. Диосгенинът се използва за лечение на остеопороза при възрастни плъхове ( 64 ). Малини посочи, че екстрактът от копър има естрогенна активност при мъжките и женските репродуктивни органели и увеличава теглото на гениталната система ( 65). Пероралното инжектиране на копър при плъхове в продължение на 10 дни увеличава теглото на млечните им жлези, яйцепровода, ендометриума, миометриума, шийките на матката и вагините. Копърът през устата увеличава концентрацията на общ протеин в семенните везикули и простатата на мъжки плъхове. Копърното масло влияе върху маточните контракции и причинява значително намаляване на интензивността на свиването, индуцирано от окситозин и Е2 простагландин ( 66 ). Това показва, че антиоксидантните съединения в копъра и витамин Е, заедно с други антиоксиданти, могат да намалят вредното въздействие на ЕМП върху репродуктивната система.
3. Заключения
Много изследвания показват, че електромагнитните полета могат да имат разрушително въздействие върху половите хормони, функцията на половите жлези, развитието на плода и бременността. Така че хората трябва да са наясно с негативните ефекти на ЕМП. Въпреки че въздействието на вълните варира при различни честоти, по-добре е да останете възможно най-далеч от техния произход поради рисковете, свързани с излагането на тези вълни. Освен това хората могат да използват естествени антиоксиданти, за да помогнат за намаляване на въздействието на тези вълни.
Благодарности
Авторът е благодарен на служителите в библиотеката на Университета по медицински науки в Тебриз.
Бележки под линия
Прожекция на iThenticate: 11 май 2016 г., редакция на английски: 02 юни 2016 г., контрол на качеството: 04 юли 2016 г.
Конфликт на интереси:
Няма конфликт на интереси, който да бъде деклариран.
Принос на авторите:
Всички автори допринесоха за този проект и статия еднакво. Всички автори прочетоха и одобриха окончателния ръкопис.
Препратки
1. Веселинова Л. Индекс на телесна маса като фактор за прогнозиране и предотвратяване на риска за професионално смесено натоварване с ниска интензивност на ЕМП. Electromagn Biol Med. 2015 г .; 34 (3): 238–43. doi: 10.3109 / 15368378.2015.1076449. [ PubMed ] [ CrossRef ] [ Google Scholar ]2. Tabrah FL, Ross P, Hoffmeier M, Gilbert F., Jr Клинично репо за дългосрочна костна плътност след краткосрочно приложение на ЕМП. Биоелектромагнетика. 1998; 19 (2): 75–8. [ PubMed ] [ Google Scholar ]3. Juutilainen J. Ефекти върху развитието на електромагнитните полета. Биоелектромагнетика. 2005: 107–15. doi: 10.1002 / bem.20125. [ PubMed ] [ CrossRef ] [ Google Scholar ]4. Gye MC, Park CJ. Ефект на електромагнитното поле, експозиция върху репродуктивната система. Clin Exp Reprod Med. 2012; 39 (1): 1–9. doi: 10.5653 / cerm.2012.39.1.1. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]5. Лондон SJ, Thomas DC, Bowman JD, Sobel E, Cheng TC, Peters JM. Излагане на битови електрически и магнитни полета и риск от детска левкемия. Am J Epidemiol. 1991; 134 (9): 923–37. [ PubMed ] [ Google Scholar ]6. Lai H, NP Singh. Индуцирана от магнитно поле ДНК верига се счупва в мозъчните клетки на плъховете. Перспектива за здравето на околната среда. 2004; 112 (6): 687–94. doi: 10.1289 / ehp.6355. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]7. Сценир. Потенциални ефекти върху здравето при излагане на електромагнитни полета. Научен комитет по възникващи и новооткрити рискове за здравето; 2015. [ PubMed ] [ Google Scholar ]8. Leszczynski D, Joenvaara S, Reivinen J, Kuokka R. Нетермично активиране на стресовия път hsp27 / p38MAPK чрез излъчване на мобилен телефон в човешки ендотелни клетки: молекулярен механизъм за ефекти, свързани с рак и кръвно-мозъчна бариера. Диференциация. 2002; 70 (2–3): 120–9. doi: 10.1046 / j.1432-0436.2002.700207.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]9. Dasdag S, Akdag MZ, Ayyildiz O, Demirtas OC, Yayla M, Sert C. Клетъчните телефони променят ли кръвните параметри и теглото при раждане на плъхове? Electromagn Biol Med. 2000; 19 (1): 107–13. doi: 10.1081 / JBC-100100301. [ CrossRef ] [ Google Scholar ]10. Luukkonen J, Hakulinen P, Maki-Paakkanen J, Juutilainen J, Naarala J. Подобряване на химически индуцираното производство на реактивни кислородни видове и увреждане на ДНК в човешки SH-SY5Y клетки на невробластома чрез радиочестотно лъчение от 872 MHz. Mutat Res. 2009; 662 (1–2): 54–8. doi: 10.1016 / j.mrfmmm.2008.12.005. [ PubMed ] [ CrossRef ] [ Google Scholar ]11. Aitken RJ, Bennetts LE, Sawyer D, Wiklendt AM, King BV. Влияние на радиочестотното електромагнитно излъчване върху целостта на ДНК в мъжката зародишна линия. Int J Androl. 2005; 28 (3): 171–9. [ PubMed ] [ Google Scholar ]12. Cao YN, Zhang Y, Liu Y. Ефекти от излагането на изключително нискочестотни електромагнитни полета върху възпроизводството на женски мишки и развитието на потомството. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi. 2006; 24 (8): 468–70. [ PubMed ] [ Google Scholar ]13. Harrington JM, McBride DI, Sorahan T, Paddle GM, van Tongeren M. Професионално излагане на магнитни полета във връзка със смъртността от мозъчен рак сред работещите в производството и преноса на електроенергия. Occup Environ Med. 1997; 54 (1): 7–13. doi: 10.1136 / oem.54.1.7. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]14. Sobel E, Dunn M, Davanipour Z, Qian Z, Chui HC. Повишен риск от болест на Алцхаймер сред работници с вероятно излагане на електромагнитно поле. Неврология. 1996; 47 (6): 1477–81. doi: 10.1212 / WNL.47.6.1477. [ PubMed ] [ CrossRef ] [ Google Scholar ]15. Brainard GC, Kavet R, Kheifets LI. Връзката между електромагнитното поле и експозицията на светлина на мелатонин и риск от рак на гърдата: преглед на съответната литература. J Pineal Res. 1999; 26 (2): 65–100. doi: 10.1111 / j.1600-079X.1999.tb00568.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]16. Mancinelli F, Caraglia M, Abbruzzese A, d'Ambrosio G, Massa R, Bismuto E. Нетермични ефекти на електромагнитните полета при честота на мобилния телефон върху повторното сгъване на вътреклетъчен протеин: миоглобин. J Cell Biochem. 2004; 93 (1): 188–96. doi: 10.1002 / jcb.20164. [ PubMed ] [ CrossRef ] [ Google Scholar ]17. Lin H, Blank M, Rossol-Haseroth K, Goodman R. Регулиране на гени с елементи на електромагнитна реакция. J Cell Biochem. 2001; 81 (1): 143–8. [ PubMed ] [ Google Scholar ]18. Heynick LN, Merritt JH. Радиочестотни полета и тератогенеза. Биоелектромагнетика. 2003: 174–86. [ PubMed ] [ Google Scholar ]19. Хавас Магда. Инженерна и технологична история. 1976. Биологични ефекти на електромагнитното лъчение. [ Google Scholar ]20. Седмици Б. Wi-Fi и неврологични заболявания. Седмици MD. 2015 г. [ Google Scholar ]21. Dimitris J, Olle J, George L. електромагнитни полета, по отношение на биологичната активност. 2015. Поляризация: Ключова разлика между изкуственото и естественото. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]22. Linda S, Amel H, Ghiraz S, Ines A, Azzouz A, Sakly M. Ефекти от острото излагане на Wi-Fi сигнали (2,45 GHz) върху вариабилността на сърцето и кръвното налягане при заек албинос. Токсикология на околната среда и фармакология. 2015 г .; 40 (2): 600–5. [ PubMed ] [ Google Scholar ]23. Hardell L, Carlberg M, Gee D. 21 Използване на мобилен телефон и риск от мозъчен тумор: ранни предупреждения, ранни действия? 2015 г. [ Google Scholar ]24. Volkow ND, Tomasi D, Wang GJ, Vaska P, Fowler JS, Telang F, et al. Ефекти от излагането на радиочестотен сигнал на клетъчния телефон върху метаболизма на глюкозата в мозъка. ДЖАМА. 2011; 305 (8): 808–13. doi: 10.1001 / jama.2011.186. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]25. Poulris AF. Ефекти върху възпроизводството и развитието на ЕМП в модели на гръбначни животни. Патофизиология. 2009; 16 (2–3): 179–89. doi: 10.1016 / j.pathophys.2009.01.010. [ PubMed ] [ CrossRef ] [ Google Scholar ]26. Nelson JF, Karelus K, Bergman MD, Felicio LS. Невроендокринно участие в стареенето: доказателства от изследвания на репродуктивното стареене и ограничаване на калориите Невробиол стареене. 1995; 16 (5): 837–43. doi: 10.1016 / 0197-4580 (95) 00072-M. [ PubMed ] [ CrossRef ] [ Google Scholar ]27. Skokri S, Soltani A, Kazemi M, Sardari D, Babapoor Mofrad F. Ефект на излагане на Wi-Fi (2,45 GHz) върху апоптозата, параметрите на сперматозоидите и хистоморфометрията на тестисите при плъхове. Cell J. 2015; 17 (2): 322–31. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]28. Kim YW, Kim HS, Lee JS, Kim YJ, Lee SK, Seo JN, et al. Ефекти от 60 Hz 14 micro T магнитно поле върху апоптозата на тестикуларната зародишна клетка при мишки. Биоелектромагнетика. 2009; 30 (1): 66–72. doi: 10.1002 / bem.20448. [ PubMed ] [ CrossRef ] [ Google Scholar ]29. Khaki AA, Zarrintan S, Khaki A, Zahedi A. Ефектите на EMF върху микроструктурата на семенните везикули при плъхове: изследване на светлинен и електронен микроскоп. Pak J Biol Sci. 2008; 11 (5): 692–701. doi: 10.3923 / pjbs.2008.692.701. [ PubMed ] [ CrossRef ] [ Google Scholar ]30. Khaki AA, Soleimanirad J, Arjani H, Mohadjel Shoja MA, Zarrintan S, Khalili A, et al. Изследване на въздействието на електромагнитните полета върху мъжкото безплодие и начините за намаляване на неговите вредни ефекти. Медицински вестник на Университета по медицински науки в Тебриз. 2006; 28 (4): 41–7. [ Google Scholar ]31. Hamdi BA, Soleimanirad J, Khiki AA, Roshangar L. Излагане на развитие на emf и неговия ефект върху сперматогенезата в зряла възраст при мишки. Международно списание за репродуктивна биомедицина. 2011; 9 (1): 67. [ Google Scholar ]32. Iorio R, Scrimaglio R, Rantucci E, DelleMonache S, Di Gaetano A, Finetti N, et al. Предварително проучване на влиянието на осцилиращото електромагнитно поле върху подвижността на човешките сперматозоиди. Био електромагнитни. 2007; 28 (1): 72–5. doi: 10.1002 / bem.20278. [ PubMed ] [ CrossRef ] [ Google Scholar ]33. Imai N, Kawabe M, Hikage T, Nojima T, Takahashi S, Shirai T. Ефекти върху тестисите на плъхове от 1.95-GHz W-CDMA за клетъчни телефони IMT-2000. Syst BiolReprod Med. 2011; 57 (4): 204–9. doi: 10.3109 / 19396368.2010.544839. [ PubMed ] [ CrossRef ] [ Google Scholar ]34. Aitken RJ, Bennetts LE, Sawyer D, Wiklendt AM, King BV. Влияние на радиочестотното електромагнитно излъчване върху целостта на ДНК в мъжката зародишна линия. Int J Androl. 2005; 28 (30): 171–9. doi: 10.1111 / j.1365-2605.2005.00531.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]35. Saygin M, Caliskan S, Karahan N, Koyu A, Gumral N, Uguz A. Тестикуларна апоптоза и хистопатологични промени, индуцирани от 2,45 GHz електромагнитно поле. ToxicolInd Health. 2011; 27 (5): 455–63. doi: 10.1177 / 0748233710389851. [ PubMed ] [ CrossRef ] [ Google Scholar ]36. Sarkar S, Ali S, Behari J. Ефект на микровълнова печка с ниска мощност върху генома на мишката: директен ДНК анализ. Mutat Res. 1994; 320 (1–2): 141–7. doi: 10.1016 / 0165-1218 (94) 90066-3. [ PubMed ] [ CrossRef ] [ Google Scholar ]37. Lee JS, Ahn SS, Jung KC, Kim YW, Lee SK. Ефекти от излагане на електромагнитно поле от 60 Hz върху апоптозата на зародишните клетки на тестисите при мишки. Азиатски J Androl. 2004; 6 (1): 29–34. [ PubMed ] [ Google Scholar ]38. Krewski D, Glickman BW, Habash RW, Habbick B, Lotz WG, Mandeville R, et al. Последни постижения в научните изследвания в областта на радиочестотите и здравето: 2001–2003. J Toxicol Environment Health B Crit Rev. 2007; 10 (4): 287–318. [ PubMed ] [ Google Scholar ]39. Roychoudhury S, Jedlicka J, Parkanyi V, Rafay J, Ondruska L, Massanyi P, et al. Влияние на 50 hz свръхнискочестотно електромагнитно поле върху подвижността и скоростта на оплождане на сперматозоидите при зайци. J Environment Sci Health A Tox Hazard Subst Environment Eng. 2009; 44 (10): 1041–7. doi: 10.1080 / 10934520902997029. [ PubMed ] [ CrossRef ] [ Google Scholar ]40. Bernabo N, Tettamanti E, Pistilli MG, Nardinocchi D, Berardinelli P, Mattioli M, et al. Ефекти от 50 Hz магнитно поле с изключително ниска честота върху морфологията и функцията на сперматозоидите от глиган, капацитирани in vitro. Териогенология. 2007; 67 (4): 801–15. doi: 10.1016 / j.theriogenology.2006.10.014. [ PubMed ] [ CrossRef ] [ Google Scholar ]41. Hamdi BA, Roshangar L, Khaki AA, Soleimani Rad J. Хистологично изследване на тестисите и параметрите на сперматозоидите при възрастни мишки, изложени на 50 Hz електромагнитно поле през периода на развитие. Анали на биологичните изследвания. 2011; 2 (5): 455–62. [ Google Scholar ]42. Burchard JF, Nguyen DH, Block E. Концентрации на прогестерон по време на естросния цикъл на млечните крави, изложени на електрическо и магнитно поле. Био Електро Магнетика. 1998; 19 (7): 438–43. doi: 10.1002 / (SICI) 1521-186X (1998) 19: 7 <438: AID-BEM6> 3.0.CO; 2-2. [ PubMed ] [ CrossRef ] [ Google Scholar ]43. Roshangar L, Hamdi BA, Khaki AA, Soleimani Rad J, Soleimani Rad S. Ефект на нискочестотното излагане на електромагнитно поле върху диференциацията на ооцитите и развитието на фоликулите. Adv Biomed Res. 2014; 3 : 76. doi: 10.4103 / 2277-9175.125874. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]44. Schnorr TM, Grajewski BA, Hornung RW, Thun MJ, Egeland GM, Murray WE, et al. Терминали за видео дисплей и риск от спонтанен аборт. N Engl J Med. 1991; 324 (11): 727–33. doi: 10.1056 / NEJM199103143241104. [ PubMed ] [ CrossRef ] [ Google Scholar ]45. Cao YN, Zhang Y, Liu Y. Ефекти от излагането на изключително нискочестотни електромагнитни полета върху възпроизводството на женски мишки и развитието на потомството. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi. 2006; 24 (8): 468–70. [ PubMed ] [ Google Scholar ]46. Goldhaber MK, Polen MR, Hiatt RA. Рискът от спонтанен аборт и вродени дефекти сред жени, които използват терминали за визуален дисплей по време на бременност. AM J IND Med. 1988; 13 : 695–706. doi: 10.1002 / ajim.4700130608. [ PubMed ] [ CrossRef ] [ Google Scholar ]47. Tenorio BM, Jimenez GC, Morais RN, Torres SM, Albuquerque Nogueira R, Silva Junior VA. Оценка на развитието на тестисите при плъхове, изложени на 60 Hz и 1 mT електромагнитно поле. J Appl Toxicol. 2011; 31 (3): 223–30. doi: 10.1002 / jat.1584. [ PubMed ] [ CrossRef ] [ Google Scholar ]48. Chung MK, Lee SJ, Kim YB, Park SC, Shin DH, Kim SH, et al. Оценка на сперматогенезата и плодовитостта при мъжки плъхове F1 след вътреутробно и новородено излагане на изключително нискочестотни електромагнитни полета. Азиатски J Androl. 2005; 7 (2): 189–94. doi: 10.1111 / j.1745-7262.2005.00007.x. [ PubMed ] [ CrossRef ] [ Google Scholar ]49. Khaki A, Ranjbar M, Rahimi F, Ghahramanian A. Ефектите на електромагнитното поле (EMF) върху яйчниците при плъхове. Ултразвук в акушерството и гинекологията. 2011; 38 : 269. doi: 10.1002 / uog.9974. [ CrossRef ] [ Google Scholar ]50. Jung KA, Ahn HS, Lee YS, Gye MC. Ефект от излагане на магнитно поле с 20 kHz на естрозния цикъл при мишки. J Microbiol Biotechnol. 2007; 17 (3): 398–402. [ PubMed ] [ Google Scholar ]51. Cecconi S, Gualtieri G, Di Bartolomeo A, Troiani G, Cifone MG, Canipari R. Оценка на ефектите от изключително нискочестотни електромагнитни полета върху развитието на фоликулите на бозайници. Hum Reprod. 2000; 15 (11): 2319–25. doi: 10.1093 / humrep / 15.11.2319. [ PubMed ] [ CrossRef ] [ Google Scholar ]52. Айдин М, Чевик А, Кандемир ФМ, Юксел М, Апайдин АМ. Оценка на хормонални промени, биохимични параметри и хистопатологичен статус на матката при плъхове, изложени на 50-Hz електромагнитно поле. Toxicol Ind Health. 2009; 25 : 153–8. doi: 10.1177 / 0748233709102717. [ PubMed ] [ CrossRef ] [ Google Scholar ]53. Yamashita H, Hata K, Yamaguchi H, Tsurita G, Wake K, Watanabe S, et al. Краткосрочното излагане на 1439-MHz TDMA сигнал не оказва естрогенен ефект при плъхове. Био Електро Магнетика. 2010; 31 (7): 573–5. doi: 10.1002 / bem.20593. [ PubMed ] [ CrossRef ] [ Google Scholar ]54. Oral B, Guney M, Ozguner F, Karahan N, Mungan T, Comlekci S, et al. Ендометриална апоптоза, предизвикана от 900-MHz мобилен телефон: превантивни ефекти на витамини Е и С. Adv Ther. 2006; 23 (6): 957–73. doi: 10.1007 / bf02850217. [ PubMed ] [ CrossRef ] [ Google Scholar ]55. Borhani N, Rajaei F, Salehi Z, Javadi A. Анализ на фрагментацията на ДНК в миши ембриони, изложени на изключително нискочестотно електромагнитно поле. Electromagn Biol Med. 2011; 30 (4): 246–52. doi: 10.3109 / 15368378.2011.589556. [ PubMed ] [ CrossRef ] [ Google Scholar ]56. Roshangar L, soleimani Rad J. Електронно микроскопско изследване на фоликулогенезата след излагане на електромагнитно поле. Списание за размножаване и безплодие. 2004; 5 (4): 299–307. [ Google Scholar ]57. soleimani Rad J, Rowshangar L, Karimi K. Ефектът на електромагнитното поле върху фалопиевата тръба. IFFS 2001 Избрана свободна комуникация, MonduzziEditore. Отдел за международни производства; Молбърн. Novamber; 2001. с. 25–30. [ Google Scholar ]58. Hajhosseini L, Khaki A, Merat E, Ainehchi N. Ефект на розмаринова киселина върху сертоли клетки Апоптоза и нива на серумни антиоксиданти при плъхове след излагане на електромагнитни полета. Afr J tradit допълнение Altern Med. 2013; 10 (6): 477–80. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]59. Bakhshaeshi M, Khaki A, Fathizad F, Khaki AA, Ghadamkheir E. Антиоксидантна роля на кверцетин, получен от Allium cepa върху алдехид оксидаза и хепатоцити апоптоза при стрептозотоцин-индуциран диабет плъх. Азиатски Pac J Trop Biomed. 2012; 2 (7): 528–31. doi: 10.1016 / S2221-1691 (12) 60090-2. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]60. Hemadi M, Saki G, Rajabzadeh A, Khodadadi A, Sarkaki A. Ефектите от прилагането на мед и витамин Е върху апоптозата при тестисите на плъхове, изложени на шумов стрес. J Hum Reprod Sci. 2013; 6 (1): 54–8. doi: 10.4103 / 0974-1208.112383. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]61. Zigo F, Farkasova Z, Elecko J, Lapin M, Chripkova M, Czerski A. Ефект на парентералното приложение на селен и витамин Е върху здравословното състояние на млечната жлеза и върху избраните антиоксидантни показатели в кръвта на млечните крави. Pol J Vet Sci. 2014; 17 (2): 217–23. doi: 10.2478 / pjvs-2014-0031. [ PubMed ] [ CrossRef ] [ Google Scholar ]62. Khazaei M, Montaseri A, Khazaei MR, Khanahmadi M. Изследване на ефекта на Foeniculumvulgare върху фоликулогенезата при женски мишки. Int J FertilSteril. 2011; 5 (3): 122–7. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]63. Kang P, Kim KY, Lee HS, Min SS, Seol GH. Противовъзпалителни ефекти на анетол при индуцирано от липополизахарид остро нараняване на белия дроб при мишки. Life Sci. 2013; 93 (24): 955–61. [ PubMed ] [ Google Scholar ]64. Chainy GB, Manna SK, Chaturvedi MM, Aggarwal BB. Анетолът блокира както ранните, така и късните клетъчни реакции, трансдуцирани от фактора на туморната некроза; ефект върху NF-KappaB, AP-1, LNK, MAPKK и апоптоза. Онкоген. 2000; 19 (25): 2943–50. doi: 10.1038 / sj.onc.1203614. [ PubMed ] [ CrossRef ] [ Google Scholar ]65. Zhao S, Nio F, Xu CY, Liu Y, Ye L, Bi GB, et al. Диосгенинът предотвратява загубата на костна маса при индуцирана от ретиноева киселина остеопороза при плъхове. Ir J Med Sci. 2015 г. doi: 10.1007 / s11845-015-1309-2. [ PubMed ] [ CrossRef ] [ Google Scholar ]66. khorshidi N. Клинични ефекти на етеричното масло от копър върху първичната дисменорея. Ирански вестник за фармацевтични изследвания. 2003: 89–93. [ Google Scholar ]Генотоксичност, предизвикана от излагане на плода и бебето на магнитни полета и модулация на ефектите на йонизиращото лъчение. Udroiu, Antoccia, et al. PLoS One (2015).
Резюме
Заден план
Малко проучвания са изследвали токсичността и генотоксичността на изключително нискочестотни магнитни полета (ELF-MF) по време на пренатално и новородено развитие. Тези фази от живота се характеризират с клетъчна пролиферация и диференциация, което може да ги направи чувствителни към стресови фактори от околната среда. Въпреки че in vitro доказателства предполагат, че ELF-MF може да модифицира ефектите на йонизиращото лъчение, досега не са провеждани изследвания in vivo за генотоксичните ефекти на ELF-MF, комбинирани с рентгенови лъчи.
Цел и методи
Целта на това проучване е да изследва в соматични и зародишни клетки ефектите от хроничната експозиция на ELF-MF от средата на бременността до отбиването и всяка възможна модулация, произведена от експозиция на ELF-MF върху увреждане, причинено от йонизиращо лъчение. Мишките бяха изложени на 50 Hz, 65 μT магнитно поле, 24 часа / ден, в продължение на общо 30 дни, започвайки от 12 дни след зачеването. Друга група е била облъчена с 1 Gy рентгенови лъчи непосредствено преди излагане на ELF-MF, други групи са били само рентгеново облъчени или изложени на измама. Микроядрен тест върху кръвни еритроцити се извършва многократно от 1 до 140 дни след раждането. Освен това, 42 дни след раждането, генотоксичните и цитотоксичните ефекти върху мъжките зародишни клетки се оценяват чрез анализ на комети и проточен цитометричен анализ.
Резултати
Излагането на ELF-MF няма тератогенен ефект и не влияе върху оцеляването, растежа и развитието. Тестът за микроядрите показа, че ELF-MF предизвиква леко генотоксично увреждане само след максималното време на експозиция и че този ефект изчезва през месеците след края на експозицията. ELF-MF няма ефект върху генотоксичността, индуцирана от йонизиращо лъчение (IR) в еритроцитите. По различен начин ELF-MF изглежда модулира отговора на мъжките зародишни клетки на рентгенови лъчи с въздействие върху процесите на пролиферация / диференциация. Тези резултати сочат значението на тъканната специфичност и развитието върху въздействието на ELF-MF върху ранните етапи от живота и показват необходимостта от допълнителни изследвания на молекулярните механизми, лежащи в основата на ELF-MF биологичните ефекти.
Фигури
Цитиране: Udroiu I, Antoccia A, Tanzarella C, Giuliani L, Pacchierotti F, Cordelli E, et al. (2015) Генотоксичност, предизвикана от излагане на плода и бебето на магнитни полета и модулация на ефектите на йонизиращото лъчение. PLoS ONE 10 (11): e0142259. https://doi.org/10.1371/journal.pone.0142259
Редактор: Мария Росария Скарфи, Национален съвет за изследвания, ИТАЛИЯ
Получено: 4 юни 2015 г .; Приет: 20 октомври 2015 г .; Публикувано: 11 ноември 2015 г.
Авторско право: © 2015 Udroiu et al. Това е статия с отворен достъп, разпространявана съгласно условията на лиценза за признание на Creative Commons , която позволява неограничено използване, разпространение и възпроизвеждане на всякакъв носител, при условие че оригиналният автор и източник са кредитирани
Наличност на данни: Всички съответни данни са в хартията и нейните поддържащи информационни файлове.
Финансиране: Тази работа е подкрепена от "Istituto nazionale per l'assicurazione contro gli infortuni sul lavoro", номер на безвъзмездна помощ B23 / DIPIA / 2009 ( www.inail.it ). LG (INAIL).
Конкуриращи се интереси: Авторите са декларирали, че не съществуват конкуриращи се интереси.
Въведение
Възможният повишен риск от рак - особено детска левкемия - свързан с изключително нискочестотни магнитни полета (ELF-MF) е причина за безпокойство [ 1 , 2 ]. Публикувани са много епидемиологични проучвания, но не е доказана недвусмислено ясна връзка между излагането на ELF-MF и рака [ 3 ]. Въпреки че IARC класифицира този физически агент като „евентуално канцерогенен за хората (група 2B)“ [ 4 ], над 1000 проведени до момента механистични проучвания все още не са разкрили възможния биологичен механизъм, чрез който ELF-MF може да причини някакъв здравен ефект [ 5]. ].
Тъй като увреждането на ДНК се счита за основната причина за рак, много изследвания изследват способността на ELF-MF да навреди на генома. Те включват голям брой изследвания, както in vitro, така и in vivo , но резултатите все още са противоречиви (както е прегледано в [ 6 ]). Сред анализите за генотоксичност тестът за микроядра е един от най-използваните, поради своята простота, чувствителност и надеждност. Този тест играе важна роля поради добрата си предсказуемост за канцерогенни опасности [ 7 ]. Досега има малко съобщения за in vivo генотоксични ефекти на ELF-MF, използващи микроядрени анализи. Svedenstal и Johanson [ 8] не открива никакви разлики в микроядрените еритроцити между възрастни мишки, изложени за 90 дни на 14 μT магнитно поле, и тези, които не са изложени; същият резултат се наблюдава от Abramsson-Zetterberg и Grawé [ 9 ], като се използва равно поле, както при възрастни, така и при новородени мишки. Обратно, бяха открити положителни резултати, като се анализират еритроцитите на новородени мишки, пренатално изложени на 650 μT [ 10 , 11 ], възрастни плъхове, изложени на 1 mT в продължение на 45 дни [ 12 ], възрастни мишки, изложени на 5 μT в продължение на 40 дни [ 13 ] и възрастни мишки изложени на 200 μT в продължение на 7 дни [ 14]. Тъй като една от основните причини за безпокойство по отношение на нейонизиращата радиация е възможната им връзка с рака в детска възраст, би било от значение да се изследват ефектите на ELF-MF по време на ранна детска възраст. Освен това, изучаването на ефектите, индуцирани от ELF-MF във фетални и неонатални етапи на живот, може да бъде полезно за разкриване на техните генотоксични свойства, тъй като бебешките клетки [ 15 - 18 ] и дори повече фетални клетки [ 19 ] показват по-голяма чувствителност към генотоксичните обиди, отколкото възрастните клетки. Доколкото ни е известно, само две работи изследват генотоксичните ефекти на ELF-MF при гризачи, изложени вътреутробно , давайки противоположни резултати [ 9 , 10 ]. Важността да се изследва вътреутробное посочена и експозицията за оценка на потенциалната канцерогенност на ELF-MF [ 20 ].
Освен това животът на плода е критична стъпка и в развитието на мъжката репродуктивна система. При гризачите първичните зародишни клетки се редуват между митотична активност и успокоение и, за разлика от възрастните, също клетките на Сертоли се размножават активно [ 21 ]. Има някои доказателства, че експозицията на ELF-MF може да повлияе на репродуктивната система на мъжете при възрастни [ 22 , 23 ], въпреки че други проучвания отчитат отрицателни резултати [ 24 ]. Съществуват малко проучвания за излагане на плода на ELF-MF на мъжката репродуктивна система. Резултатите от мултигенерационно проучване при плъхове не подкрепят хипотезата за репродуктивен или токсичен ефект върху развитието [ 25 ] и не се наблюдават промени в сперматогенезата и плодовитостта на потомството при плъхове следпри вътреутробна и неонатална експозиция [ 26 ]. От друга страна, McGivern и колеги [ 27 ] наблюдават увеличаване на теглото при епидидимус, простата и семенни мехурчета при възрастни плъхове, изложени вътреутробно . Промяна на сперматогенен епител също се наблюдава при възрастни плъхове след вътреутробна експозиция [ 28 ] и след вътреутробна / неонатална експозиция [ 29 ].
Въпреки че има някои доказателства, че ELF-MF може да повлияе на реакцията на увреждане на ДНК, предизвикана от IR in vitro [ 30 - 32 ], не е провеждано проучване in vivo за възможните генотоксични ефекти от комбинирано излагане на ELF-MF и X- лъчи.
Следователно, в тази работа ние имаме за цел да проучим ефектите от ниско ниво на хронична експозиция на ELF-MF при мишки по време на много чувствителен период като фетален и неонатален живот и всяка възможна модулация, която експозицията на ELF-MF може да окаже върху уврежданията, причинени IR. Освен това, тъй като отдавна е доказано [ 33 ], че IR може да доведе до забавени ефекти ( de novo ефекти при необлъчени потомци или съседи на облъчени клетки), но няма данни за влиянието, което комбинираното излагане на множество физически агенти може да окаже върху забавени ефекти, ние също изследвахме резултатите от комбинираните експозиции върху геномната нестабилност.
Материали и методи
Животни и система за експозиция
Бременни CD-1 швейцарски (безпородни) мишки (река Чарлз, Италия) бяха разделени на четири групи, включващи по два язовира. Една група е била неекспонирана и е служила за контрол (С, 27 малки); друга група (E, 20 малки) е била изложена на ELF-MF от ден 11.5 след зачеването (pc) до отбиването, общо 30 дни; друга група (X, 25 малки) е била облъчена с X [1 Gy) на ден 11,5 pc; последната група (XE, 31 малки) е била облъчена с X (1 Gy) на ден 11,5 pc и незабавно изложена на ELF магнитни полета до отбиването (общо 30 дни).
За този експеримент използвахме магнитно поле от 65 μT. Тъй като мишките се нуждаят от 12-26 пъти по-голяма експозиция на MF от тази, изисквана от хората, за да предизвикат подобна плътност на тока в тялото [ 34 ], използваното от нас поле е (по отношение на биологичните ефекти) сравним с 2–5 μT MF за хората. Тези стойности обикновено присъстват в повечето домакинства.
Магнитното поле с 50 Hz, 65 μT се генерира от соленоид, работещ 24 часа на ден. Соленоидът беше с дължина 0,8 m и радиус 0,13 m, с 552 оборота от 2,5 mm 2 медна тел, навита на два слоя непрекъснато напред-назад около цилиндър от PVC. Той се захранва от 50 Hz основно захранване чрез трансформатор. Приложено е напрежение от 6,5 V (rms), за да се получи плътност на потока от 65 μT (rms) в центъра на соленоида. Полето е равномерно между ± 5% от обема, където са били изложени мишките. Соленоидът не е екраниран за електрическото поле, тъй като индуцираното електрическо поле е незначително поради използваното ниско напрежение. Излагането на 1 Gy рентгенови лъчи се извършва в апарат Gilardoni (Gilardoni, Италия) при скорост на дозата 0,5 Gy / min.
Животните бяха настанени в поликарбонатни клетки, поставени вътре в работещ соленоид (групи E и XE) или изключен соленоид (групи C и X). Температурата и относителната влажност в помещението за животни са съответно 22 ° C и 40%. Изкуственото осветление беше от 8 сутринта до 8 вечерта и търговските пелети и чешмяна вода бяха на разположение ad libitum през целия експериментален период. Температурата в бобините беше същата като в стаята.
Размерът на котилото се определя при раждането. Малките се претеглят при раждането и на 11, 21, 42 и 140 ден след раждането. Степента на оцеляване се оценява при отбиването (ден 21). Ежедневно се наблюдават малките за поява на физиологични забележителности на развитие.
Четиридесет и три мъжки мишки бяха умъртвени на 42 дни след раждането за анализи върху репродуктивната система. Останалите животни бяха жертвани на 140 ден след раждането. Жертвата е извършена чрез дислокация на шийката на матката при упоени животни.
Експериментът е проведен съгласно италианските закони, регулиращи използването и хуманното отношение към животните за научни цели (указ № 116/1992). Всички експериментални процедури са одобрени от Етичния комитет за изследване на животните към италианското министерство на здравеопазването (ID на одобрение: 10.10.15).
Микроядрен тест върху еритроцити
Получени са кръвни мазки, пробиващи опашната вена на мишките при раждането и на 11-ия ден (кърмаческа възраст), 21 (отбиване), 42 (полова зрялост) и 140 (зряла възраст) след раждането. Плъзгачите се фиксират в абсолютен метанол и се поддържат при -20 ° С до оцветяване. Смазите се оцветяват с акридинов портокал (Sigma-Aldrich, Италия, 20 μg / ml в рН 6.8 буфер Sørensen). Пробите бяха кодирани и отбелязани сляпо от същия анализатор. Микроядрата се оценяват при увеличение 1000 пъти, като се използва флуоресцентен микроскоп Zeiss Axiophot (Zeiss, Германия) (възбуждащ филтър 494 nm, бариерен филтър 523 nm). За всяко животно бяха анализирани 2000 еритроцити [ 10 ].
Анализ на комета върху епидидимална сперма
След жертва, както епидидима caudae са хирургично отстранени, се поставя в блюдо на Петри, съдържаща TNE буфер (0.15 М NaCl, 0.01 М Tris-HCl, 0.001 М Na 2 EDTA, рН 7.4) и смляно с извити ножици. Суспензиите на сперматозоиди се филтрират през 0,2 mm найлонова мрежа, центрофугират се (5 минути, 1800 g), ресуспендират се в TNE буфер плюс 10% глицерол, аликвотират и се замразяват при -80 ° С. Алкални (рН 12,1) и неутрални (рН 8,0) протоколи от анализа на Комета бяха извършени за оценка на увреждането на ДНК в сперматозоидите. Анализът се извършва по същество според Cordelli et al. [ 35 ]. Накратко стъкла се потапят в разтвор на лизиране (2.5 М NaCl, 100 тМ Na 2EDTA, 10 mM Tris, рН 10,0), съдържащ 10% DMSO (Carlo Erba, Италия) и 1% Triton X-100 (Sigma-Aldrich), една нощ при 4 ° C. В края на лизиса предметните стъкла се потапят за 30 минути в 10 mM дитиотреитол (Sigma-Aldrich) в лизисен разтвор, за да декондензират изключително уплътнения сперматозоиден хроматин и да позволят миграцията на ДНК. След това предметните стъкла бяха поставени в хоризонтален резервоар за електрофореза с гел, където електрофорезата беше извършена при следните условия: 10 минути при 4 ° С в алкален буфер за електрофореза (300 mM NaOH, 1 mM Na 2 EDTA; HCl беше добавена за достигане на рН 12.1), последвано от чрез 7 минути 27 V (0,8 V / cm), 300 mA електрофореза, при 4 ° С, за алкалния анализ; 20 минути в ТВЕ буфер (2 мМ Na 2EDTA, 90 mM Tris, 90 mM борна киселина; рН 8,0) при 4 ° C, последвано от 7 минути 27 V (0,8 V / cm), 10 mA електрофореза, при 4 ° C, за неутралния анализ. След електрофореза предметните стъкла се фиксират за 5 минути в Tris 0,4 М рН 7,5 и за 5 минути в абсолютен етанол и се сушат на въздух при стайна температура. Непосредствено преди точкуването, предметните стъкла се оцветяват с 12 μg / ml етидиев бромид (Sigma-Aldrich) и се изследват при увеличение 200 пъти с флуоресцентен микроскоп на Olympus. Слайдовете бяха анализирани сляпо с компютърна система за анализ на изображения (Delta Sistemi, Италия). За да се оцени количеството на увреждане на ДНК, бяха анализирани 100 клетки от два различни предмета и беше използван компютърно генериран процент на стойностите на ДНК на опашката (интензитет на опашката).
Поточен цитометричен анализ на разпределението на субпопулациите на тестисите
След отстраняване на tunica albuginea, тестисите се смилат с хирургически ножици и се обработват с 0,1% разтвор на пепсин (1,5 милионсона единици / mg; Серва, Германия) в продължение на 10 минути при стайна температура при магнитно разбъркване, за да се увеличи максимално освобождаването на зародишните клетки от семенните каналчета. Клетъчната суспензия се фиксира в 70% студен етанол. След това пробите се съхраняват при -20 ° С преди анализ. Фиксираните клетки се третират с 0,5% разтвор на пепсин в продължение на 10 минути при стайна температура при магнитно разбъркване и се оцветяват с разтвор, съдържащ 50 μg / ml пропидиев йодид (Sigma-Aldrich) и 40 μg / ml RNase (Sigma-Aldrich), в PBS. Клетките, оцветени с пропидиев йодид, бяха анализирани в FACSCalibur поточен цитометър (Becton-Dickinson Immunocytometry, CA, USA). Записани са флуоресцентните сигнали на оцветени с пропидиев йодид клетки, и цитограма на съдържанието на ДНК спрямо броя на клетките се използва за идентифициране на клетъчни популации въз основа на тяхното съдържание на ДНК. Типичните хистограми за разпределение на интензивността на флуоресценцията на ДНК от клетки на тестисите на възрастни мишки се характеризират с различни пикове, включващи клетки с различно съдържание на ДНК. Първият регион (1С) включва постмейотични хаплоидни сперматиди. Диплоидна G1-фазна сперматогония и вторични сперматоцити се записват във втория пик (2С), заедно с крайно диференцирани тестисни соматични клетки. Третият връх (4C) включва някои G2 / M сперматогонии, но най-вече първични сперматоцити. Активно синтезиращите ДНК клетки от S-фаза са разположени в областта между втория и третия пик. Типичните хистограми за разпределение на интензивността на флуоресценцията на ДНК от клетки на тестисите на възрастни мишки се характеризират с различни пикове, включващи клетки с различно съдържание на ДНК. Първият регион (1С) включва постмейотични хаплоидни сперматиди. Диплоидна G1-фазна сперматогония и вторични сперматоцити се записват във втория пик (2С), заедно с терминално диференцирани тестисни соматични клетки. Третият пик (4C) включва някои G2 / M сперматогонии, но най-вече първични сперматоцити. Активно синтезиращите ДНК клетки от S-фаза са разположени в областта между втория и третия пик. Типичните хистограми за разпределение на интензивността на флуоресценцията на ДНК от клетки на тестисите на възрастни мишки се характеризират с различни пикове, включващи клетки с различно съдържание на ДНК. Първият регион (1С) включва постмейотични хаплоидни сперматиди. Диплоидна G1-фазна сперматогония и вторични сперматоцити се записват във втория пик (2С), заедно с терминално диференцирани тестисни соматични клетки. Третият пик (4C) включва някои G2 / M сперматогонии, но най-вече първични сперматоцити. Активно синтезиращите ДНК клетки от S-фаза са разположени в областта между втория и третия пик. и вторичните сперматоцити се записват във втория пик (2С), заедно с крайно диференцирани тестисни соматични клетки. Третият пик (4C) включва някои G2 / M сперматогонии, но най-вече първични сперматоцити. Активно синтезиращите ДНК клетки от S-фаза са разположени в областта между втория и третия пик. и вторичните сперматоцити се записват във втория пик (2С), заедно с крайно диференцирани тестисни соматични клетки. Третият връх (4C) включва някои G2 / M сперматогонии, но най-вече първични сперматоцити. Активно синтезиращите ДНК клетки от S-фаза са разположени в областта между втория и третия пик.
За всяка хистограма са записани общо 10 000 събития. Относителните честоти на типовете клетки на тестисите бяха изчислени с помощта на софтуера Cell Quest.
Статистически анализ
Разликите в оцеляването между различните групи бяха анализирани чрез процедурата на Mantel за log-rank тест. Бяха извършени множество линейни регресии, за да се оцени дали теглото и честотата на микроядрата зависят от пола, размера на постелята и лечението. Тъй като не бяха открити разлики (с помощта на t-критерия на Student) между двете котила от една и съща група, беше изчислена средна стойност спрямо всички малки, третирани по един и същи начин и използвана за сравнение между групите. T-тестът на Welch е използван за сравнение на микроядрените честоти на различните групи на лечение. Еднопосочният тест ANOVA и Дънкан е използван за post-hoc сравнение на параметрите на мъжките зародишни клетки и данните от кометен анализ между различните групи на лечение. Нивото на значимост е установено при p <0,05. Всички тестове бяха проведени с помощта на Statistica 10 (Statsoft., Inc., ОК, САЩ).
Резултати
Раждане и развитие
Таблица 1 показва размера на постелята, теглото при раждането и при отбиването и оцеляването при отбиването на четирите експериментални групи. Нито магнитните полета, нито рентгеновите лъчи изглеждаха повлияли размера на котилото. Оцеляването не се различава значително между групите. Множественият регресионен анализ показа, че ефектите от пола и размера на постелята върху теглото са по-големи от ефекта от лечението.
Микроядрен тест върху еритроцити
Множествен регресионен анализ показа, че честотите на микроядрата не са свързани значително нито с пола, нито с размера на котилото. Не са открити значителни разлики между средните микроядрени честоти на двете котила от една и съща третирана група.
Фигура 1 показва средната честота на микронуклеираните еритроцити в кръвта, взета проба по различно време в четирите групи. При раждането разликите в честотите на микроядрата (MN) са статистически значими между групи С и Х и между групи С и ХЕ ( р <0,001, и двете). По същия начин, на 11-ия ден разликите в честотите на MN между групите C и X и между групите C и XE също бяха статистически значими ( p = 0,002 и p = 0,0057, съответно). На 21-ия ден средните честоти на MN в групите C, E, X и XE не се различават значително. На 42-ия ден средните честоти на MN в E, X и XE са значително различни в сравнение с C (съответно p = 0,008, p = 0,02 ир <0,001). На ден 140 групата XE показа MN честота, статистически по-висока от тази на контролната група ( p = 0,0048). Група Х също е по-висока от С, но разликите не са значими поради по-голяма стандартна грешка (по това време броят на животните е значително намален). Трябва също така да се отбележи, че в група X (както и група XE), честотата на микроядрените еритроцити не се различава между ден 42 и 140. Разликата между групи X и XE никога не е била статистически значима.
Фигура 1. Честоти на микроядрата в еритроцитите на периферната кръв.Броят на животните във всяка група се отчита в колоните на хистограмата. Баровете представляват стандартна грешка. Значимост в сравнение с C: * p <0,05; ** р <0,01; *** р <0,001.
Ефекти върху мъжката репродуктивна система
За анализиране и на ефектите върху мъжката полова система, всички животни, принадлежащи към 4 експериментални групи бяха умъртвени 42 дни след раждането. Въздействието на експозицията (ите) се оценява от способността на системата да произвежда окончателно диференцирани зародишни клетки и нивото на увреждане в тяхната ДНК. Пренаталното облъчване с единична доза рентгенови лъчи по време на ранната диференциация на половите жлези индуцира значително намаляване на теглото на тестисите, предполагащо токсичен ефект ( Фигура 2А). Излагането на ELF-MF от 11,5 дни след зачеването до 21 дни след раждането индуцира леко намаляване на относителното тегло на тестисите, главно поради малко увеличение на телесното тегло. След комбинирано излагане на йонизиращо лъчение и ELF-MF, нивото на намаляване на теглото на тестисите е сравним с нивото, предизвикано от рентгеновите лъчи.
Фигура 2. (А) Относително тегло на тестисите (тегло на тестисите / телесно тегло * 100); (Б) проценти на 1С пост мейотични клетки, оценени чрез цитометричен анализ на ДНК съдържание на поточни клетки на цели тестиси; (C) брой на сперматозоидите в епидидима на кауда.Колоните представляват средните стойности на 7 контроли (C), 11 ELF-MF изложени (E), 14 рентгенови лъчи (X) или 11 животни, изложени на комбинирано лечение (XE). Звездичките отбелязват статистически различни стойности между групите, *: p <0,05; **: p <0,005.
Чрез поточен цитометричен анализ на тестикуларни клетки се изчисляват относителните проценти на 1С, 2С, S-фаза и 4С клетки. След експозиция на рентгенови лъчи се наблюдава значително намаление от 28% на постмейотичните 1С клетки и малко, но значително намаление от 13% се открива и след излагане на ELF-MF ( Фигура 2В ). Изненадващо, след комбинираната експозиция, процентът на 1С клетки е значително по-висок, отколкото след единичната обработка и равен на този на контролите. Доказателствата за индуцирана от рентгеновите лъчи токсичност бяха увеличени от анализа на терминално диференцирани клетки в епидидима на кауда, където беше открито намаляване на броя на сперматозоидите с 73% ( Фигура 2С). Излагането на ELF-MF не предизвиква значителна промяна в броя на сперматозоидите по отношение на контролите. В съответствие с данните за относителния процент на тестикуларните 1С клетки, броят на епидидималните сперматозоиди след комбинирана експозиция е значително по-висок, отколкото само след рентгенови лъчи и не се различава от неекспонираните контроли. Данните за теглото на епидидима са в съответствие с тези за броя на сперматозоидите (данните не са показани).
Ефектите от еднократно и комбинирано излагане на ELF-MF и рентгенови лъчи бяха допълнително оценени чрез анализ на комети в епидидимни сперматозоиди. Не е показан ефект от еднократна обработка с ELF-MF или рентгенови лъчи както при неутрални, така и при алкални условия ( Фигура 3А). Също така комбинираната експозиция не предизвиква значителни вариации по отношение на контролите, но когато данните се сравняват с тези, събрани само след рентгенови лъчи, се установява статистически значимо намаляване на средната интензивност на опашката. При неутрални условия беше установена произволна точка на прекъсване на интензивността на опашката, над която се смяташе, че сперматозоидите са ясно повредени. В съответствие с данните за средната интензивност на опашката, процентите на ясно увредени сперматозоиди не се различават значително от контролите при каквито и да било условия на лечение, но процентът на ясно увредени сперматозоиди, индуцирани от комбинираната експозиция, е значително по-малък от този, предизвикан само от рентгенови лъчи ( Фигура 3В ).
Фигура 3. Анализ на комета при епидидимни сперматозоиди.(А) Средни стойности на интензитета на опашката, получени с анализ на алкална и неутрална комета. (Б) Процент на сперматозоидите, носещи ясно увредена ДНК (със стойности на интензитета на опашката по-високи от 10%) след неутрален анализ. Колоните представляват средните стойности на 4 контроли (C), 5 ELF-MF изложени (E), 8 рентгенови лъчи (X) или 6 животни, изложени на комбинирано лечение (XE).
Дискусия
Основната цел на това проучване беше да се използва чувствителен модел за откриване на генотоксичните свойства на ELF-MF, тъй като след десетилетия проучвания доказателствата все още са противоречиви. Следователно, ние изследвахме генотоксичните ефекти на ELF-MF in vivo върху соматични и зародишни клетки след излагане по време на пренатални и неонатални етапи на развитие, за които е известно, че са чувствителни към стреса в околната среда. В допълнение, генетичните промени, възникващи през този период от живота, могат да представляват съ-фактор в началото на детския рак [ 37 , 38 ]. Насочихме се и към разследване in vivoвъзможно влияние на ELF-MF върху клетъчната реакция на йонизиращо лъчение. Хроничното излагане на ELF-MF от 12-ия ден до отбиването няма тератогенен ефект и не влияе върху оцеляването, растежа и развитието. Рентгеновото облъчване на 12-ия ден доведе до закърнели малки и забавено развитие, в съгласие с литературните данни [ 36 ]. Не са наблюдавани синергични ефекти на ELF-MF и излагане на рентгенови лъчи върху тези параметри.
Освен тези крайни точки за развитие, ние изследвахме генотоксичните ефекти върху еритроидната и мъжката зародишна линия, използвайки техники, валидирани в съответните тъкани.
Резултатите от микроядрения тест в периферна кръв изглежда показват, че излагането на ELF-MF предизвиква лека генотоксична обида, която се открива само по време на ограничен период на експресия, след максималното време на експозиция (един месец, включително пренатална и новородена Период). Този резултат е различен, но не е в противоречие с резултата, получен от Abramsson-Zetterberg и Grawé [ 9 ], които продължават експозицията по време на бременност, спират я при раждане и вземат проби от кръв от пренатално изложени мишки на възраст от 35 дни, без да се установи увеличаване на MN честотите. Всъщност разликите в продължителността и прозорците на експозиция на развитие могат да обяснят различните резултати.
Наблюдаваният слаб генотоксичен ефект на ELF-MF, наблюдаван само 42 дни след раждането, може да се дължи на нарушаване на клетъчната защитна способност срещу ендогенни източници на ДНК лезии. : изглеждаше необходимо непрекъснато излагане от средата на бременността до отбиването, последвано от период на „латентност“, с по-нататъшно развитие на хематопоетичната система. След 4 месеца системата изглежда възстанови хомеостатичния си капацитет. Трябва да се проведат специфични експерименти за идентифициране на молекулярните механизми, водещи до генотоксични увреждания. Нарушаването на окислителния баланс може да бъде кандидат, тъй като много автори предполагат, че този механизъм е замесен в ELF-MF-индуцирани биологични ефекти [ 39 , 40 ].
По различен начин, излагането на рентгенови лъчи изглежда води до два различни периода на откриваеми генотоксични ефекти. Повишена честота на MN се наблюдава при раждането като пряк ефект от рентгеновото облъчване. По времето, когато това се случи (ден 12 pc), феталният черен дроб започва еритропоеза благодарение на наскоро пристигнали хематопоетични стволови клетки [ 41 ] и микронуклеирани еритроцити, получени от тези предшественици / прекурсори, са открити в циркулиращата кръв при раждането. В съответствие с кинетиката на пренаталната еритропоеза, честотата на микронуклеираните еритроцити, наблюдавани в кръвта, е с един порядък по-малка от тази, открита във феталния хематопоетичен черен дроб по време на максимална експресия (16–18 часа) след сравнима радиационна обработка [ 42]. С течение на времето тези микроядрени клетки се разреждат допълнително в кръвния поток от еритроцити, образувани от необлъчени предшественици. Всъщност честотата на MN на X групата на 11-ия ден е по-ниска от тази при раждането и достига контролната стойност на 21-ия ден. Освен този остър, ранен изход, това, което изглежда като забавен ефект, се открива на 42-ия ден и продължава до деня 140. И накрая, данните показват, че излагането на ELF-MF не е променило ефекта на рентгеновите лъчи в нито един тестван момент.
Ефектите върху мъжката репродуктивна система са оценени 42 дни след раждането, когато всички субпопулации на зародишни клетки присъстват в тестисите, спермиогенезата е започнала и първата вълна от сперматозоиди е налична в епидидимуса. На тази възраст обаче зародишната тъкан на мишката все още е в процес на развитие, както се вижда от няколко сравнителни данни: теглото на тестисите е малко по-ниско от това на възрастните (0,11 срещу 0,14 g съответно) и броят на сперматозоидите е по-малък от 15% от този открити при възрастни мишки.
Пренаталното излагане на рентгенови лъчи на ден 11,5 pc е оказало влияние върху развитието на тестисите, както се вижда от намаляването на теглото на тестисите, на относителната честота на пост-мейотичните клетки (1С) и на броя на епидидималните сперматозоиди на 42 дни след раждането. Тези резултати са в съгласие с известната радиочувствителност на зародишни и соматични клетки във феталните полови жлези [ 43 ].
Излагането на ELF-MF от 11,5 дни след зачеването до 21 дни след раждането предизвиква леко относително намаляване на пост-мейотичните клетки, което не се отразява в намаляване на сперматозоидите. Проучванията върху ефекта на вътреутробната експозиция на ELF-MF върху репродуктивната система на мъжете са малко и резултатите са противоречиви. При възрастни плъхове се наблюдава дезорганизация на сперматогенен епител в резултат на оксидативен стрес, предизвикан от вътреутробно излагане на ELF-MF, 3 mT, 4 h / ден [ 28 ]. Промяна на тестисите се наблюдава и при плъхове, изложени от 13 дни pc до 21 дни след раждането на ELF-MF 1 mT, 3 приложения по 30 минути на ден [ 29]. Увеличение на теглото в епидидимуса, простатата и семенните везикули, без разлики в броя на епидидималните сперматозоиди, се наблюдава при възрастни плъхове, изложени in utero на импулсно електромагнитно поле от 15 Hz от 15 до 20 ден на бременността [ 27 ]. От друга страна, вътреутробното и новородено излагане на плъхове на ELF-MF при полеви сили до 500 μT не води до промяна в сперматогенезата и плодовитостта на потомството [ 26 ]. Не са показани токсични ефекти върху репродукцията или развитието при многогенерационно проучване при плъхове, изложени на 18,5 часа / ден на ELF-MF при полеви сили до 1 mT [ 25 ].
Доколкото ни е известно, не са провеждани проучвания за оценка на взаимодействието на рентгеновите лъчи и ELF-MF преди раждането при бозайниците. Пафкова и колеги [ 44] съобщава, че 10 mT магнитно поле променя ембриотоксичния ефект на йонизиращото лъчение върху пилешки ембриони. Нашите резултати предполагат, че непрекъснатата експозиция на ELF-MF от 11,5 дни pc, непосредствено след облъчване с рентгенови лъчи, до 21 дни след раждането, модулира токсичния ефект на йонизиращото лъчение, както е показано от по-високия процент на пост-мейотичните тестикуларни клетки и броя на епидидимни сперматозоиди в XE по отношение на X експериментална група. Нашият експериментален протокол не е насочен към изучаване на възможните ранни механизми в основата на този ефект, но могат да бъдат изведени няколко хипотези: ELF-MF може да повлияе реакцията на увреждане на ДНК, стимулирайки възстановяването на ДНК или инхибирайки апоптозата; освен това може да се предвиди въздействие на ELF-MF върху пролиферацията на клетки, оцелели при рентгеновите лъчи, което също може да повлияе на настъпването на пубертета.
В нашето проучване ние също имаме за цел да оценим възможните забавени ефекти върху ДНК на терминално диференцирани сперматозоиди. Индукцията на забавени ефекти в мъжките зародишни клетки се подкрепя от наблюдението на увреждане на ДНК в сперматозоидите, произтичащо от рентгенови лъчи, изложени на възрастни сперматогонии [ 35 , 45 - 47 ]. В настоящото проучване се наблюдава малко, незначително увеличение на увреждането на ДНК след вътреутробноизлагане на 1 Gy рентгенови лъчи. Трябва да се отбележи, че нивото на увреждане на ДНК в сперматозоидите на нелекувани 42-дневни мишки е било по-високо и по-хетерогенно, отколкото при възрастни мишки (лабораторни исторически данни не са показани). Този резултат е в съгласие с характеристиките на популациите на сперматозоиди, произведени от първата вълна на сперматогенеза, които в сравнение с възрастните сперматозоиди показват аномалии в морфологията на сперматозоидите и анормалната структура на хроматина [ 48]. Не са открити ефекти след продължително излагане на 50 Hz, 65 μT ELF-MF. Изненадващо се наблюдава очевидно намаляване на увреждането на ДНК, хомогенно при всички животни от групата, когато рентгеновите лъчи са последвани от ELF-MF експозиция. Този ефект, заедно с наблюдаваното увеличаване на броя на сперматозоидите, предполага, че ELF-MF е в състояние да модулира отговора на мъжките зародишни клетки на рентгеновите лъчи. Това откритие може да се дължи на влияние на ELF-MF върху ранната реакция на увреждане на ДНК, водещо до намаляване на рентгеновите убийства на клетки. Нещо повече, ELF-MF би могъл да повлияе на пътищата за пролиферация / диференциация на зародишни клетки, вероятно предвиждайки началото на сперматогенезата в пубертета; това би довело до разреждане на първата вълна от естествено възникващи анормални сперматозоиди с непроменени сперматозоиди, получени от втора вълна на сперматогенеза.4 , 49 ].
Интензивността на магнитното поле, което използвахме, беше доста висока; въпреки това, той е в порядъка на степента на някакво битово облъчване, например в случая на държане на преносим компютър в утробата. В зародишната тъкан успяхме да открием биологичен ефект на ELF-MF, особено като модулация на клетъчния отговор на токсичната инсулт, предизвикана от единична доза рентгенови лъчи. Този резултат потвърждава важността да се изследват ефектите от комбинираното излагане на ELF-MF и други стресови фактори, тъй като модулацията на реакцията на стрес може да бъде толкова критична за клетъчната съдба, колкото и самият стрес. Нашите данни подкрепят хипотезата, че пластичността на развиващите се тъкани ги прави уязвими към външни стресови фактори. Освен това, ако ELF-MF наистина може да повлияе на процесите на пролиферация / диференциация през ранните фази от живота, важно е да се изследват различни тъкани / органи, тъй като всеки от тях има свой специфичен модел на диференциация в определен времеви прозорец на развитие. Това от своя страна сочи към необходимостта от по-задълбочено разбиране на молекулярните механизми, залегнали в основата на ELF-MF биологичните ефекти.
подкрепяща информация
S1 данни. Индивидуални стойности за микроядрения тест (Dataset1) и за мъжката репродуктивна система (Dataset2).
https://doi.org/10.1371/journal.pone.0142259.s001
(PDF)
Благодарности
Авторите са благодарни на A. Bedini, C. Giliberti и R. Palomba за техническата помощ при излагане на магнитно поле на ELF.
Принос на автора
Замисля и проектира експериментите: CT AA AS IU. Изпълнени експерименти: EC PV PE IU. Анализирани данни: FP EC PV PE IU AS. Реактиви / материали / инструменти за анализ, допринесени: FP EC PV PE AS LG. Написа хартията: FP EC PV IU AS.
Препратки
- 1.Wertheimer N, Leeper E. Конфигурации на електрически кабели и рак на детството. Am J Epidemiol. 1979; 109 (3): 273–284. pmid: 453167
- 2.Teepen JC, ван Dijck JA. Влияние на високите нива на електромагнитно поле върху честотата на детската левкемия. Int J Рак. 2012; 131 (4): 769–778. pmid: 22437882
- 3.Repacholi M. Загриженост, че магнитните полета "EMF" от електропроводи причиняват рак. Sci Total Environment. 2012; 426: 454–458. pmid: 22534362
- 4.Работна група на IARC за оценка на канцерогенните рискове за хората. Нейонизиращо лъчение, Част 1: статични и изключително нискочестотни (ELF) електрически и магнитни полета. IARC Monogr Eval Канцерогенни рискове Hum. 2002; 80: 1–395. pmid: 12071196
- 5.Световна здравна организация. Критерии за екологично здраве 238, Изключително нискочестотни полета; 2007 г.
- 6.Udroiu I, Giuliani L, Ieradi LA. Генотоксични свойства на изключително нискочестотни електромагнитни полета. В: Giuliani L, Soffritti M, редактори. Нетермични ефекти и механизми на взаимодействие между електромагнитните полета и живата материя. Фиденца (Италия): Матиоли 1885; 2010. стр.123–134.
- 7.Morita T, Asano N, Awogi T, Sasaki YF, Sato S, Shimada H, et al. Оценка на микроядрен анализ на гризачи при скрининг на IARC канцерогени (групи 1, 2А и 2В) обобщения доклад на 6-то съвместно проучване от CSGMT / JEMS MMS. Съвместно проучване на теста за микроядрена група. Изследователска група за мутагенност при бозайници. Mutat Res. 1997; 389 (1): 3–122. pmid: 9062586
- 8.Svedenstal BM, Johanson KJ. Левкоцити и микронуклеирани еритроцити в периферна кръв от мишки, изложени на 50-Hz или 20-kHz магнитни полета. Електро-магнитобиол. 1998; 17: 127–143.
- 9.Abramsson-Zetterberg L, Grawé J. Продължителното излагане на възрастни и фетални мишки на 50 Hz магнитно поле не увеличава честотата на микроядрата в еритроцитите. Биоелектромагнетика. 2001; 22 (5): 351–357. pmid: 11424159
- 10.Udroiu I, Cristaldi M, Ieradi LA, Bedini A, Giuliani L, Tanzarella C. Кластогенност и анеуплоидия при новородени и възрастни мишки, изложени на 50 Hz магнитни полета. Int J Radiat Biol. 2006; 82 (8): 561–567. pmid: 16966183
- 11.Udroiu I, Cristaldi M, Ieradi LA, Bedini A, Giuliani L. Генотоксични и хематоксични увреждания, индуцирани от ELF магнитни полета. Eur J Oncol. 2008; 13: 239–244.
- 12.Erdal N, Gürgül S, Celik A. Цитогенетични ефекти на изключително нискочестотно магнитно поле върху костен мозък на плъх Wistar. Mutat Res. 2007; 630 (1–2): 69–77. pmid: 17452120
- 13.Okudan N, Celik I, Salbacak A, Cicekcibasi AE, Buyukmumcu M, Gökbel H. Ефекти от дългосрочно излагане на магнитно поле от 50 Hz върху честотата на микроядрените полихроматични еритроцити и кръвни лимфоцити и аргирофилните ядрени организаторни области в лимфоцитите на мишки. Neurol Endocrinol Lett. 2010; 31: 208–214.
- 14.Alcaraz M, Olmos E, Alcaraz-Saura M, Achel DG, Castillo J. Ефект от дългосрочното излагане на магнитно поле от 50 Hz върху микроядрените полихроматични еритроцити на мишки. Electromagn Biol Med. 2014; 33 (1): 51–57. pmid: 23781994
- 15.Sanderson BJ, Clark AM. Микроядра при възрастни и фетални мишки, изложени in vivo на хелиотрин, уретан, монокроталин и бензидин. Mutat Res. 1993; 285 (1): 27–33. pmid: 7678129
- 16.Udroiu I, Ieradi LA, Cristaldi M, Tanzarella C. Откриване на кластогенни и анеугенни увреждания при новородени плъхове. Environment Mol Mutagen. 2006; 47 (5): 320–324. pmid: 16538686
- 17.Zúñiga-González G, Ramírez-Muñoz MP, Torres-Bugarín O, Pérez-Jiménez J, Ramos-Mora A, Zamora-Pérez A, Gallegos-Arreola MP, Sánchez-Corona J. Индукция на микронуклеуси в домашната котка (Felis domesticus ) периферна кръв от колхицин и цитозин-арабинозид. Mutat Res. 1998; 413 (2): 187–189. pmid: 9639702
- 18.Neri M, Ceppi M, Knudsen LE, Merlo DF, Barale R, Puntoni R, Bonassi S. Базова честота на микроядрите при деца: оценки от мета- и обединени анализи. Environmental Health Perspect. 2005; 113 (9): 1226–1229. pmid: 16140632
- 19.Paashuis-Lew YR, Heddle JA. Спонтанна мутация по време на развитието на плода и постнаталния растеж. Мутагенеза. 1998; 13 (6): 613–617. pmid: 9862193
- 20.Li P, McLaughlin J, Infante-Rivard C. Професионално излагане на майката на изключително нискочестотни магнитни полета и риск от рак на мозъка при потомството. Ракът причинява контрол. 2009; 20 (6): 945–955. pmid: 19224378
- 21.Vergouwen RP, Jacobs SG, Huiskamp R, Davids JA, de Rooij DG. Пролиферативна активност на гоноцити, клетки на Sertoli и интерстициални клетки по време на развитието на тестисите при мишки. J Reprod Fertil. 1991; 93 (1): 233–243. pmid: 1920294
- 22.Chernoff N, Rogers JM, Kavet R. Преглед на литературата за потенциална репродуктивна токсичност и токсичност за развитието на електрически и магнитни полета. Токсикология. 1992; 74 (2–3): 91–126. pmid: 1519247
- 23.Lee SK, Park S, Gimm YM, Kim YW. Изключително нискочестотните магнитни полета предизвикват апоптоза на сперматогенни зародишни клетки: възможен механизъм. Biomed Res Int. 2014; 2014: 567183. pmid: 25025060
- 24.Duan W, Liu C, Wu H, Chen C, Zhang T, Gao P, Luo X, Yu Z, Zhou Z. Ефекти от излагането на изключително нискочестотни магнитни полета върху сперматогенезата при възрастни плъхове. Биоелектромагнетика. 2014; 35 (1): 58–69. pmid: 24122970
- 25.Райън BM, Symanski RR, Pomeranz LE, Johnson TR, Gauger JR, McCormick DL. Оценка на репродуктивна токсичност на много поколения на магнитни полета от 60 Hz, използвайки непрекъснат протокол за размножаване при плъхове. Тератология. 1999; 59 (3): 156–162. pmid: 10194806
- 26.Chung MK, Lee SJ, Kim YB, Park SC, Shin DH, Kim SH, Kim JC. Оценка на сперматогенезата и плодовитостта при мъжки плъхове F1 след вътреутробно и новородено излагане на изключително нискочестотни електромагнитни полета Азиатски J Androl. 2005; 7 (2): 189–194. pmid: 15897976
- 27.McGivern RF, Sokol RZ, Adey WR. Пренаталното излагане на нискочестотно електромагнитно поле демаскулинизира поведението на маркиращия аромат на възрастни и увеличава теглото на допълнителните полови органи при плъхове. Тератология. 1990; 41 (1): 1–8. pmid: 2106174
- 28.Gharamaleki H, Parivar K, Soleimani Rad J, Roshangar L, Shariati M. Ефекти от експозицията на електромагнитно поле през пренаталния период върху биомаркери на оксидативен стрес и патология на тестисите и нивото на тестостерон при възрастни плъхове във F1 поколение. Акта Ендо. (Buc.) 2014; 10: 577–587.
- 29.Tenorio BM, Jimenez GC, Morais RN, Torres SM, Albuquerque Nogueira R, Silva Junior VA. Оценка на развитието на тестисите при плъхове, изложени на 60 Hz и 1 mT електромагнитно поле. J Appl Toxicol. 2011; 31 (3): 223–230. pmid: 20936650
- 30.Hintenlang DE. Синергични ефекти на йонизиращото лъчение и 60 Hz магнитни полета. Биоелектромагнетика. 1993; 14 (6): 545–551. pmid: 8297398
- 31.Miyakoshi J, Yoshida M, Shibuya K, Hiraoka M. Излагането на силни магнитни полета при честота на мощността потенцира рентгенови индуцирани разкъсвания на ДНК вериги. J Radiat Res. 2000; 41 (3): 293–302. pmid: 11210830
- 32.Yoon HE, Lee JS, Myung SH, Lee YS. Повишен γ-H2AX чрез излагане на магнитни полета от 60 Hz, комбинирани с йонизиращо лъчение, но не и водороден прекис, в нетуморогенни човешки клетъчни линии. Int J Radiat Biol. 2014; 90 (4): 291–298. pmid: 24467330
- 33.Karotki AV, Baverstock K. Какви механизми / процеси стоят в основата на радиационно-индуцирана геномна нестабилност? Cell Mol Life Sci. 2012; 69 (20): 3351–60. pmid: 22955377
- 34.Xi W, Stuchly MA, Gandji OP. Индуцирани електрически токове в модели на човек и гризачи от магнитни полета от 60 Hz. IEEE TransBiomed Eng. 1994; 41: 1018–1023.
- 35.Cordelli E, Fresegna AM, Leter G, Eleuteri P, Spanò M, Villani P. Оценка на увреждане на ДНК в различни етапи на сперматогенеза на мишки след облъчване на тестисите X. Radiat Res. 2003; 160 (4): 443–451. pmid: 12968930
- 36.Brent RL. Лъчева тератогенеза. Тератология. 1980; 21 (3): 281–298. pmid: 7006137
- 37.Маршал GM, Carter DR, Cheung BB, Liu T, Mateos MK, Meyerowitz JG, Weiss WA. Пренаталният произход на рака. Nat Rev Рак. 2014; 14 (4): 277–289. pmid: 24599217
- 38.Kim AS, Eastmond DA, Preston RJ. Детска остра лимфоцитна левкемия и перспективи за оценка на риска от експозиция в ранен стадий. Mutat Res. 2006; 613 (2–3): 138–160. pmid: 17049456
- 39.Luukkonen J, Liimatainen A, Juutilainen J, Naarala J. Индукция на геномна нестабилност, окислителни процеси и митохондриална активност от 50 Hz магнитни полета в човешки SH-SY5Y клетки на невробластома. Mutat Res. 2014; 760: 33–41. pmid: 24374227
- 40.Mattsson MO, Simkó M. Групиране на експериментални условия като подход за оценка на ефектите от изключително нискочестотни магнитни полета върху окислителния отговор при in vitro проучвания. Front Public Health. 2014; 2: 132. pmid: 25229055
- 41.Ema H, Nakauchi H. Разширяване на хемопоетични стволови клетки в развиващия се черен дроб на миши ембрион. Кръв. 2000; 95 (7): 2284–2288. pmid: 10733497
- 42.Cole RJ, Taylor N, Cole J, Arlett CF. Краткосрочни тестове за трансплацентарно активни канцерогени. I. Образуване на микроядра във фетални и майчини миши еритробласти. Mutat Res. 1981; 80 (1): 141–157. pmid: 7207479
- 43.Vergouwen RP, Huiskamp R, Bas RJ, Roepers-Gajadien HL, Davids JA, de Rooij DG. Радиочувствителност на тестикуларните клетки при феталната мишка. Radiat Res. 1995; 141 (1): 66–73. pmid: 7997516
- 44.Pafkova H, Jerabek J. Взаимодействие на MF 50 Hz, 10 mT с висока доза рентгенови лъчи: оценка на ембриотоксичността при ембриони на пилета. Rev Environment Health. 1994; 10 (3–4): 235–241. pmid: 7724884
- 45.Haines GA, Hendry JH, Daniel CP, Morris ID. Повишени нива на кометно открито увреждане на ДНК сперматозоиди след in vivo изотопно или рентгеново облъчване на сперматогонии. Mutat Res. 2001; 495 (1–2): 21–32. pmid: 11448639
- 46.Haines GA, Hendry JH, Daniel CP, Morris ID. Увреждане на зародишните клетки и зависимото от дозата ДНК, измерено чрез анализа на кометите в миши сперматозоиди след рентгеново облъчване на тестисите Biol Reprod. 2002; 67 (3): 854–861. pmid: 12193394
- 47.Cordelli E, Eleuteri P, Grollino MG, Benassi B, Blandino G, Bartoleschi C, Pardini MC, Di Caprio EV, Spanò M, Pacchierotti F, Villani P. Директно и забавено рентгеново индуцирано увреждане на ДНК в мъжки зародишни клетки на мишки. Environment Mol Mutagen. 2012; 53 (6): 429–439. pmid: 22730201
- 48.Janca FC, Jost LK, Evenson DP. Развитие на тестисите и сперматозоидите на мишки, характеризиращо се от раждането до зряла възраст чрез двойна параметрична поточна цитометрия Biol Reprod. 1986; 34 (4): 613–623. pmid: 3708046
- 49.Zhang X, Zhang J, Qu X, Wen J. Ефекти на различни изключително нискочестотни електромагнитни полета върху остеобластите. Electromagn Biol Med. 2007; 26 (3): 167–177. pmid: 17886004
- Йон Удроиу ,
- Антонио Анточа,
- Патриция Елевтери,
- Паола Вилани,
- Антонела Сгура
- Публикувано: 11 ноември 2015 г.
- https://doi.org/10.1371/journal.pone.0142259
Оксидативният стрес на мозъка и черния дроб се увеличава чрез Wi-Fi (2,45 GHz) излагане на плъхове по време на бременност и развитие на новородени. Likelik, Ömer, et al. Journal of Chemical Neuroanatomy 75 (B): 134-139 (2015).
Оксидативният стрес на мозъка и черния дроб се увеличава чрез Wi-Fi (2,45 GHz) излагане на плъхове по време на бременност и развитието на новородени
Принадлежности- PMID: 26520617
- DOI: 10.1016 / j.jchemneu.2015.10.005
Резюме
Прекомерното производство на реактивни кислородни вещества (ROS) и намалените антиоксидантни защитни системи в резултат на излагане на електромагнитна радиация (EMR) може да доведе до окислително увреждане на мозъка и черния дроб и разграждане на мембраните по време на бременност и развитие на малки плъхове. Целта ни беше да изследваме ефектите на индуцираната от Wi-Fi EMR върху мозъчно-чернодробните антиоксидантни редокс системи при плъхове по време на бременност и развитие. Шестнадесет бременни плъхове и техните 48 новородени са разделени по равно на контролни и EMR групи. EMR групите бяха изложени на 2,45 GHz EMR (1 h / ден в продължение на 5 дни / седмица) от бременността до 3-седмична възраст. Взети са проби от мозъчна кора и черен дроб от новородените между първата и третата седмица. В EMR групите нивата на липидна пероксидация в мозъка и черния дроб бяха повишени след излагане на EMR; въпреки това, активността на глутатион пероксидазата (GSH-Px) и концентрациите на витамин А, витамин Е и β-каротин са намалени в мозъка и черния дроб. Концентрациите на глутатион (GSH) и витамин С в мозъка също са по-ниски в EMR групите, отколкото в контролите; концентрациите им обаче не се променят в черния дроб. В заключение, индуцираният от Wi-Fi оксидативен стрес в мозъка и черния дроб на развиващите се плъхове е резултат от намалените концентрации на GSH-Px, GSH и антиоксидантните витамини. Освен това мозъкът изглежда е по-чувствителен към окислително увреждане в сравнение с черния дроб при развитието на новородени. Концентрациите на глутатион (GSH) и витамин С в мозъка също са по-ниски в EMR групите, отколкото в контролите; концентрациите им обаче не се променят в черния дроб. В заключение, индуцираният от Wi-Fi оксидативен стрес в мозъка и черния дроб на развиващите се плъхове е резултат от намалените концентрации на GSH-Px, GSH и антиоксидантните витамини. Освен това мозъкът изглежда е по-чувствителен към окислително увреждане в сравнение с черния дроб при развитието на новородени. Концентрациите на глутатион (GSH) и витамин С в мозъка също са по-ниски в EMR групите, отколкото в контролите; концентрациите им обаче не се променят в черния дроб. В заключение, индуцираният от Wi-Fi оксидативен стрес в мозъка и черния дроб на развиващите се плъхове е резултат от намалените концентрации на GSH-Px, GSH и антиоксидантните витамини. Освен това мозъкът изглежда е по-чувствителен към окислително увреждане в сравнение с черния дроб при развитието на новородени.
Ключови думи: Антиоксидантни витамини; Мозък; Електромагнитно излъчване; Глутатион; Черен дроб; Оксидативен стрес.
Copyright © 2015 Elsevier BV Всички права запазени.
Подобни статии
- Запасите от чернодробни антиоксиданти предпазват мозъка от индуциран от електромагнитна радиация (900 и 1800 MHz) оксидативен стрес при плъхове по време на бременност и развитието на потомство.J Matern Fetal Neonatal Med. 2014 декември; 27 (18): 1915-21. doi: 10.3109 / 14767058.2014.898056. Epub 2014 9 април.PMID: 24580725
- Дългосрочното излагане на електромагнитно излъчване от мобилни телефони и Wi-Fi устройства намалява плазмените нива на пролактин, прогестерон и естроген, но увеличава оксидативния стрес на матката при бременни плъхове и техните потомци.Ендокринни. 2016 май; 52 (2): 352-62. doi: 10.1007 / s12020-015-0795-3. Epub 2015 14 ноември.PMID: 26578367
- Wi-Fi (2,45 GHz) - и мобилен телефон (900 и 1800 MHz) - индуцирани рискове от оксидативен стрес и елементи в бъбреците и тестисите на плъхове по време на бременност и развитието на потомство.Biol Trace Elem Res. 2013 декември; 156 (1-3): 221-9. doi: 10.1007 / s12011-013-9836-z. Epub 2013 8 октомври.PMID: 24101576
- Последни доклади за Wi-Fi и индуцирано от мобилен телефон лъчение на оксидативен стрес и репродуктивни сигнални пътища при жени и мъже.J Membr Biol. 2013 декември; 246 (12): 869-75. doi: 10.1007 / s00232-013-9597-9. Epub 2013 9 октомври.PMID: 24105626 Преглед.
- [Влияние на електромагнитното излъчване от мобилни телефони и Wi-Fi върху сперматогенезата].Zhonghua Nan Ke Xue. 2019 май; 25 (5): 451-455.PMID: 32216233 Преглед. Китайски.
Цитирано от 8 статии
- Wi-Fi (2,4 GHz) засяга антиоксидантния капацитет, експресията на гени за възстановяване на ДНК и апоптозата при плацентата на бременна мишка.Iran J Basic Med Sci. 2020 юни; 23 (6): 833-840. doi: 10.22038 / ijbms.2020.40184.9512.PMID: 32695301 Безплатна статия от PMC.
- Доказателства за оксидативен стрес след продължително излагане на Wi-Fi лъчение при модел на плъх.Environment Sci Pollut Res Int. 2018 декември; 25 (35): 35396-35403. doi: 10.1007 / s11356-018-3482-0. Epub 2018 21 октомври.PMID: 30343375
- Скептични подходи относно ефекта от излагането на електромагнитни полета върху мозъчните хормони и ензимните дейности.J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 177-184. doi: 10.1016 / j.jmau.2017.09.002. Epub 2017 14 септември.PMID: 30023252 Безплатна статия от PMC. Преглед.
- Умерена доза Trolox, предотвратяваща вредните ефекти на Wi-Fi радиацията върху сперматозоидите in vitro чрез намаляване на уврежданията от оксидативен стрес.Chin Med J (англ.). 2018 20 февруари; 131 (4): 402-412. doi: 10.4103 / 0366-6999.225045.PMID: 29451144 Безплатна статия от PMC.
- Промяна на адаптивното поведение на потомството след излагане на майчин мобилен телефон.Environment Sci Pollut Res Int. 2018 април; 25 (11): 10894-10903. doi: 10.1007 / s11356-017-1178-5. Epub 2018 3 февруари.PMID: 29397508
Невродегенеративни промени и апоптоза, предизвикани от вътрематочно и извънматочно излагане на радиочестотна радиация. Güler, Göknur, et al. Journal of Chemical Neuroanatomy 75 (B): 128-133 (2015). J Chem Neuroanat
doi: 10.1016 / j.jchemneu.2015.10.006. Epub 2015 28 октомври.. 2016 г. септември; 75 (Pt B): 128-33.Невродегенеративни промени и апоптоза, предизвикани от вътрематочно и извънматочно излагане на радиочестотна радиация
Принадлежности- PMID: 26520616
- DOI: 10.1016 / j.jchemneu.2015.10.006
Резюме
Неблагоприятните ефекти върху здравето на радиочестотната радиация (RFR) върху текущите етапи на развитие на децата от зачеването до детството са научно очаквани теми. Това проучване беше проведено, за да се идентифицират ефектите на глобалната система за мобилни комуникации (GSM), модулиран мобилен телефон като RFR в 1800MHz честота върху окислително увреждане на ДНК и липидна пероксидация до образуването на апоптотични клетки, като се използват хистопатологични и имунохистохимични методи в мозъчната тъкан на 1- едномесечни мъжки и женски новозеландски бели зайци, които са били изложени на тези полета в утробата на майка си и след раждането. Окислителното увреждане на ДНК и нивата на липидна пероксидация бяха изследвани чрез измерване на нивата на 8-хидрокси-2'-дезоксигуанозин (8-OHdG) и нива на малондиалдехид (MDA), съответно. Наблюдавани са хистопатологични промени, използвани чрез оцветяване с хематоксилин и еозин (HE). Апоптотични клетки бяха открити в изследваните органи чрез терминално дезоксинуклеотидил трансфераза-медиирано dUTP никелно маркиране (TUNEL). Както за мъжки, така и за женски бебета; Нивата на 8-OHdG се повишават в групата, изложена на RFR, както във вътрематочния, така и в извънматочния период, в сравнение с бебетата, които никога не са били изложени на RFR, и тези, които са били изложени, когато са навършили един месец (p <0,05). Резултатите от MDA са различни при мъжки и женски зайци. Няма разлика между всички женски бебешки групи (p> 0,05), докато само вътрематочната експозиция значително води до повишаване на нивото на MDA за мъжките бебета. HE оцветяването разкрива леки намаления при невронална некробиоза в мозъчните тъкани на женски зайци, които са били само с вътрематочна експозиция, а мъжките зайци са имали само извънматочна експозиция. Глиозата е леко положителна в мозъчните тъкани на зайци, които са изложени само на вътрематочен период, а групата е изложена както на вътрематочен, така и на извънматочен период. Не е установена обаче апоптотична промяна чрез оцветяване с TUNEL в мозъчните тъкани на всички групи.
Ключови думи: 8-OhdG; Апоптоза; Увреждане на ДНК; Липидна пероксидация; Мобилен телефон; ТУНЕЛ.
Copyright © 2015 Elsevier BV Всички права запазени.
Подобни статии
- Ефектът на радиочестотното лъчение върху увреждането на ДНК и липидите при женски и мъжки новородени зайци.Int J Radiat Biol. 2012 април; 88 (4): 367-73. doi: 10.3109 / 09553002.2012.646349.PMID: 22145622
- Влиянието на 1800 MHz GSM-подобни сигнали върху химията на кръвта и оксидативния стрес при небременни и бременни зайци.Int J Radiat Biol. 2012 май; 88 (5): 414-9. doi: 10.3109 / 09553002.2012.661517. Epub 2012 2 март.PMID: 22280439
- Ефекти от пренаталното и постнаталното излагане на GSM-подобна радиочестота върху химията на кръвта и оксидативния стрес при бебета зайци, експериментално проучване.Cell Biochem Biophys. 2013 ноември; 67 (2): 743-51. doi: 10.1007 / s12013-013-9564-1.PMID: 23526187
- Ефект на радиочестотната радиация върху репродуктивното здраве.Indian J Med Res. 2018 декември; 148 (Suppl): S92-S99. doi: 10.4103 / ijmr.IJMR_1056_18.PMID: 30964085 Безплатна статия от PMC. Преглед.
- Неврологични ефекти на радиочестотното лъчение.Occup Med (Lond). 2003 г.; 53 (2): 123-7. doi: 10.1093 / occmed / kqg030.PMID: 12637597 Преглед.
Излагането на майката на непрекъснато 900-MHz електромагнитно поле провокира невронална загуба и патологични промени в церебелума на 32-дневно женско потомство на плъхове. Odaci, Ersan, et al. Journal of Chemical Neuroanatomy 75 (B): 105-110 (2015).Chem Neuroanat
doi: 10.1016 / j.jchemneu.2015.09.002. Epub 2015 21 септември.. 2016 г. септември; 75 (Pt B): 105-10.Излагането на майката на непрекъснато електромагнитно поле от 900 MHz провокира невронална загуба и патологични промени в малкия мозък на 32-дневно женско потомство
Принадлежности- PMID: 26391347
- DOI: 10.1016 / j.jchemneu.2015.09.002
Резюме
Голям брой хора са несъзнателно изложени на електромагнитни полета (ЕМП) от безжични устройства. Съществуват доказателства за променено развитие на малкия мозък във връзка с пренатално излагане на ЕМП. Въпреки това все още не е налице достатъчно информация относно ефектите от излагането на 900 мегагерца (MHz) ЕМП през пренаталния период върху последващото постнатално развитие на малкия мозък. Това проучване е планирано да изследва 32-дневната женска малкия мозък на плъх след излагане на 900MHz EMF по време на пренаталния период, използвайки стереологични и хистопатологични методи за оценка. Бременните плъхове бяха разделени на контролни, фалшиви и EMF групи. Бременни EMF плъхове (PEMFG) са били изложени на 900MHz EMF в продължение на 1 час в EMF клетка по време на 13-21 дни от бременността. Бременни плъхове с фалшива група (PSG) също бяха поставени в клетката на ЕМП по време на 13 часа от бременността в продължение на 1 час, но не бяха изложени на никаква ЕМП. Не е извършена процедура при бременни плъхове от контролна група (PCG). Плъхове от контролна група за новородени (CG) са получени от PCG майки, плъхове за новородени фиктивни групи (SG) от PSG и плъхове за новородени EMF от групата (EMFG) от плъхове PEMFG. Мозъчните мозъци на новородените женски плъхове бяха извлечени на постнаталния ден 32. Броят на клетките на Purkinje беше оценен стереологично, а хистопатологичните оценки бяха извършени и на малките мозъчни срезове. Общият брой на клетките на Purkinje, изчислен чрез стереологичен анализ, е значително по-нисък в EMFG в сравнение с CG (p <0,05) и SG (p <0,05). Освен това, някои патологични промени като пикнотични неврони с тъмна цитоплазма са наблюдавани в EMFG секции под светлинна микроскопия. В заключение, резултатите от нашето проучване показват, че пренаталното излагане на ЕМП влияе върху развитието на клетките на Purkinje в малкия мозък на женски плъх и че последствията от този патологичен ефект продължават да съществуват и след постнаталния период.
Ключови думи: малкия мозък; Електромагнитно поле; Женски плъх; Клетка Пуркине; Стереология.
Copyright © 2015 Elsevier BV Всички права запазени.
Подобни статии
- Броят на клетките на Purkinje намалява при малкия мозък на възрастни женски плъхове след излагане на 900 MHz електромагнитно поле.Brain Res. 2010 г. 14 октомври; 1356: 95-101. doi: 10.1016 / j.brainres.2010.07.103. Epub 2010 4 август.PMID: 20691167
- Дългосрочното излагане на непрекъснато електромагнитно поле от 900 MHz нарушава мозъчната морфология при млади възрастни мъжки плъхове.Biotech Histochem. 2017; 92 (5): 324-330. doi: 10.1080 / 10520295.2017.1310295. Epub 2017 16 май.PMID: 28506085
- Морфологични и антиоксидантни увреждания в гръбначния мозък на мъжки потомци плъхове след излагане на непрекъснато 900MHz електромагнитно поле през ранната и средата на юношеството.J Chem Neuroanat. 2016 септември; 75 (Pt B): 99-104. doi: 10.1016 / j.jchemneu.2015.11.006. Epub 2015 17 дек.PMID: 26708410
- Изпитване на електромагнитни полета за потенциална канцерогенна активност: критичен преглед на животински модели.Environmental Health Perspect. 1997 февруари; 105 Suppl 1 (Suppl 1): 81-103. doi: 10.1289 / ehp.97105s181.PMID: 9114279 Безплатна статия от PMC. Преглед.
- Въздействия върху пренаталното развитие на човешкия мозък: систематичен преглед.J Matern Fetal Neonatal Med. 2017 октомври; 30 (20): 2461-2468. doi: 10.1080 / 14767058.2016.1253060. Epub 2016 22 ноември.PMID: 27806674 Преглед.
Цитирано от 4 статии
- Излагането на електромагнитно поле по време на бременността влияе неблагоприятно на електрофизиологичните свойства на клетките на Purkinje при потомство на плъхове.J Biomed Phys Eng. 2020 г. 1 август; 10 (4): 433-440. doi: 10.31661 / jbpe.v0i0.560. eCollection 2020 августPMID: 32802791 Безплатна статия от PMC.
- Ефект на 900-MHz електромагнитно поле върху малкия мозък: Хистопатологично изследване.Sisli Etfal Hastan Tip Bul. 2018 юни 1; 52 (2): 129-134. doi: 10.14744 / SEMB.2018.42275. eCollection 2018.PMID: 32595386 Безплатна статия от PMC.
- Импулсните мощни микровълни не нарушават функциите на кожните нормални и раковите клетки in vitro : Краткосрочна биологична оценка.J Adv Res. 2019 20 ноември; 22: 47-55. doi: 10.1016 / j.jare.2019.11.007. eCollection 2020 мартPMID: 31956441 Безплатна статия от PMC.
- Радиочестотни електромагнитни радиационни промени в поведението и тяхната възможна основа.Environment Sci Pollut Res Int. 2019 октомври; 26 (30): 30693-30710. doi: 10.1007 / s11356-019-06278-5. Epub 2019 28 август.
Различни периоди на вътрематочна експозиция на електромагнитно поле: Влияние върху плодовитостта на женските плъхове, пренаталното и постнаталното развитие. Alchalabi, Aklilu, et al. Азиатско-тихоокеански вестник за възпроизвеждане 5 (1): 14-23 (2015).
Оригинално изследванеРазлични периоди на вътрематочно излагане на електромагнитно поле: Влияние върху плодовитостта на женските плъхове, пренаталното и постнаталното развитиеПод Creative Commons лицензсвободен достъпКлючови думи
1 800 MHz GSM честотаПред- и постнатално развитиеПлъхове1 . Въведение
Потенциалният риск от радиочестотна електромагнитна вълна (RF-EMW), излъчвана от съвременните мобилни телефонни комуникационни технологии за човешката среда, и здравето е строго свързан със съвременните подходи за определяне на ограничения за безопасност за мобилни телефони, безжична точност (Wi-Fi), както и всички електронни устройства работят в обхвата на микровълните. Граници на безопасност, определени от насоките на Международната комисия за защита от нейонизиращо лъчение (ICNIRP), базирани на термичния ефект на краткосрочното излагане на нейонизиращо лъчение в микровълновия диапазон върху биологичната система [1] . В днешно време огромно количество изследователски статии от епидемиологични, човешки, животински, клетъчни, механизми и дозиметрияпроучванията сочат към нетермичен ефект на микровълновата печка (MW) върху биологичната система след хронично излагане на околната среда и човешкото население на RF-EMW (интензивно използване на мобилни телефонни комуникационни устройства) [2] , [3] , [4] , [5] . Най-новата програма за изследване на Световната здравна организация (СЗО) за радиочестотните полета (RF) подтикна проучвания върху различни възрасти на децата в допълнение към работата с животни, за да се изследва ефектът на радиочестотната експозиция върху пренаталното развитие и поведение и разглеждане на ефекта на RF върху ранните етапи [6] . Електромагнитни полета (ЕМП) с честота 900–1 800 MHz глобална система за мобилни телекомуникации (GSM) и 2 450 MHz Wi-Fi сигнали повишиха общественото безпокойство по отношение на здравния ефект с изключително внимание към подрастващото население. ICNIRP и Институтът по електротехника и електроника (IEEE) не включват бременни жени и техните бебета в техните гранични нива на безопасност на излагане на RF-EMW, въпреки че те включват работници и широка общественост в техните гранични нива на RF-EMW [1] , [ 7] . СЗО наскоро съобщи, че „натрупаните доказателства не установяват съществуването на неблагоприятни краткосрочни или дългосрочни ефекти върху здравето от сигналите, произведени от базови станции и локални безжични мрежи“ и „механистичното разбиране би разгледало възможността децата да реагират на RF повече ефективно от възрастните “[8] . През последните 10 години бяха публикувани няколко статии, показващи ефекта на вътрематочното излагане на радиочестотното излъчване (RFR) върху майките и техните новородени. Тези изследвания изследват ефекта на RFR върху оксидативен стрес , увреждане на ДНК, тератогенност , невроразвитие , ембриогенеза и поведение при лабораторни животни и птичи видове [5] , [9] , [10] , [11] , [12] . Те предполагат, че RFR и GSM-подобни сигнали са имали неблагоприятни ефекти върху бременните язовири, както и върху техните новородени и са имали вредни ефекти върху птичи ембриони след излагане на мобилен телефон или генератор на сигнали с честота от 850 MHz – 1 800 MHz. От друга страна, няколко отчитат противоречиви резултати по отношение на ефекта от вътрематочната Wi-Fi експозиция на бременни плъхове и постнаталното развитие . Проучванията за тератология и развитие не са открили никакви вредни ефекти от излагането на радиочестотни полета, свързани с мобилен телефон, при нива на експозиция под стандартните нива [13] . Противоречиво проучване в Турция установи ограничаване на растежа и забавен пубертет при женски плъхове Wister поради вътрематочна и ранна експозиция на Wi-Fi сигнал [2]. Дългосрочното излагане на „интензивна употреба“ на мобилни телефонни комуникационни технологии или чрез мобилни телефони, или чрез излъчване на базови станции на мобилни телефони, което е основният и важен източник на RFR в нашата среда по време на различни етапи на бременността, трябва да се разглежда като сериозен проблем, който трябва да бъдат адресирани адекватно. Липсата на солидно и непоколебимо убедително проучване, което установява вредния ефект на електромагнитното излъчване от мобилни телефони и базови станции върху бременни жени и новородени, обосновава необходимостта от по-нататъшни и обширни проучвания в това отношение. Следователно, това проучване е предназначено да изследва биоефектите на 1 800 MHz подобни на GSM-RF-EMF на мобилен телефон върху различни периоди на бременност върху плодовитостта на женските плъхове, пренаталното и постнаталното развитие на малките.
2 . материали и методи
2.1 . Животни и дизайн на изследване
Изследването е одобрено от научния комитет на Факултет по ветеринарна медицина на Университета в Малайзия Kelantan (UMK) и е проведено в съответствие с насоките на UMK за експерименти с животни (FPV-PGSC-2014). Плъховете Sprague – Dawley са отглеждани при стайна температура (24 ± 10) ° C и влажност от (60 ± 10)% (относителна влажност) с цикъл светлина / тъмнина 12–12 h в лабораторното звено за изследване на животни на Факултет по ветеринарна медицина, (UMK), чешмяна вода и стандартни гранули от плъхове са предоставени ad libitum . За вътрематочна експозиция през различни периоди периодът на бременността е разделен на три интервала 1-ва седмица, 2-ра седмица и 3-та седмица.
Използвани са шейсет девствени женски плъхове (във всеки интервал) на възраст 12 седмици в началото на експеримента. Женските са били чифтосвани с неекспонирани възрастни плодови мъжки плъхове от същия щам в съотношение 1: 2 (♂: ♀) за максимум 15 нощи. По време на периода на съжителство, вагинални проби се изследват микроскопски всяка сутрин за наличие на сперма. Денят на намиране на копулаторна запушалка или сперма се счита за гестационен ден 0. Всички жени се наблюдават ежедневно за смъртност и за физически признаци от началото на експозицията. Животните се държат в клетки от плексиглас. По време на експозицията се използва специално проектирана експозиционна кутия от плексиглас (60 cm × 40 cm × 20 cm), тъй като плексигласът е непроводим материал, който не се влияе от RF-EMR. Протоколът за излагане на RF-EMR беше направен, както следва:
- а)
Излагането на интервал от 1-ва седмица започва от Gestation Day (GD) 0 до GD 7.
- б)
Излагането на интервал от 2-ра седмица започва от GD 0 до GD 14.
- ° С)
Излагането на интервал от 3-та седмица започва от GD 0 до GD20 един ден преди раждането.
Всеки експериментален интервал се състои от:
- 1)
Контролна група (n = 20).
- 2)
Група за експозиция 1 h / ден (n = 20).
- 3)
Група за експозиция 2 h / ден (n = 20).
В края на всеки интервал от експеримента експерименталните животни бяха разделени на две подгрупи; А и Б. Плодовитостта на жените и пренаталното развитие се оценяват във всеки интервал от женските А на подгрупа, докато постнаталното развитие се оценява при малките от женските В подгрупа.
2.2 . Настройка на радиочестотното електромагнитно излъчване
Бременните язовири бяха разпределени на случаен принцип в девет групи (три групи / интервал и 20 животни / група). Нивото на SAR 0,048 W / Kg беше изчислено с помощта на уравнението,
Където (E) е величината на електрическото поле 38,63 v / m, (σ) е проводимостта 1,34 s / m, а (ρ) е масовата плътност на тъканната еквивалентна среда 1 090 kg / m 3 [14] . Бяха направени експозиции на радиочестотно излъчване на цялото тяло (RFR). ( Фигура 1 ).

Фигура 1 . Интервал 2-ра седмица от бременността (A) женска жена показва нормална матка с нормално разпределение на ембрионите в двата маточни рога , (B, C) маточните рога на жените от двете групи на експозиция показват задръствания, кръвоизливи, мъртви и реабсорбирани ембриони и са показани със стрелки , (D, E и F) маточните рога на жените от двете групи на експозиция показват необичайно разпределение и намаляване на броя на ембрионите.
GSM-подобни сигнали на честота 1 800 MHz се осигуряват от генератор на сигнали (Agilent Technologies E8267D, 250 KHz – 20 GHz PSG Vector Signal Generator) с интегриран модул за импулсна модулация и антена с клаксон (A-INFOMW Standard Gain Horn Antenna 1.7 –2,6 GHz WR430, Китай) в експозиционна стая. Сигналите са модулирани амплитудно от правоъгълни импулси с честота на повторение 217 Hz и работен цикъл 1: 8 (широчина на импулса 0,576 msec), съответстващ на доминиращия модулационен компонент на GSM. RFR генераторът осигурява 20 dBm (0,1 W) мощности по време на периода на експозиция [15] , [16] , [17] .
2.3 . Наблюдения на майката
2.3.1 . Тегло на тялото
Всички животни се претеглят индивидуално на нулево време (един ден преди започване на експеримента) и в края на всеки интервал от експеримента, като се използва везна с чувствителност 0,01 g.
2.3.2 . Клинично наблюдение
Ежедневно клинично изследване се извършва на жени по време на експериментални периоди за клинични признаци: аборт, състояние на лигавицата , поведение и нервни признаци.
2.4 . Фертилитет и пренатално развитие
В края на периода на експозиция на всеки интервал, животни в подгрупи А, упоени с хлороформ и всички женски са евтаназирани на GD7, 14 и 20, направено е цезарово сечение . Матката беше отстранена, отворена и регистриран статус на бременността. По време на интервала от 1-ва седмица се изследва матката на всеки язовир, за да се определи броят на местата за имплантиране и броят на бластоцистите във всеки маточен рог . През интервала от 2-ра седмица бяха изследвани маточните рога, за да се определи (1) броят на живите или мъртвите плодовена маточен рог, (2) броят на местата за ранна и късна резорбция на всеки маточен рог. Докато траеше 3-та седмица от бременността при GD 20, общият брой на плодовете, броят на живите, мъртвите и плодовете бяха претеглени индивидуално. Макроскопските аномалии бяха проверени внимателно.
2.5 . Постнатално развитие
Жените от подгрупа Б след приключване на излагането на RF-EMR и през трите интервала от бременността са наблюдавани внимателно, докато не се случи естествено раждане . Бременните жени са разделени индивидуално в клетки за наблюдение от деня на раждането до 30 дни.
2.5.1 . Данни за доставка
Всеки язовир беше изследван след 24-ия час на естествено раждане, за да се преброи общият брой на малките / котилото / група / интервал, живи, мъртви и с аномалии.
2.5.2 . Телесно тегло на малките
Интервалите на малките / котилото / групата / бяха претеглени на 24 часа след раждането, D7, D14, D21 и D28 поотделно (взети са 30 малки / група за седмично наддаване на тегло).
2.5.3 . Нормално физиологично развитие
След общ преглед, броят на малките / котилото / групата / интервалът за наблюдение за следното физиологично развитие. 30 малки / група / интервал бяха проверени внимателно и индивидуално за следните параметри: Външен вид, външен вид на косата, външен вид на зъбите, отваряне на очите и ден на отбиване като индикатор за нормален растеж.
2.6 . Биохимичен анализ
Направен е бърз метод за измерване на малоновия диалдехид (MDA) като биомаркер за липидна пероксидация в плазмени проби, който се основава на теста за тиобарбитурова киселина (TBA) чрез използване на спектрофотометрично количествено определяне на розов комплекс MDA-TBA, образуван чрез реакция на MDA с две молекули TBA. Използвани са комплекти за анализ на липидна пероксидация (MDA) (ab118970, abcam) в съответствие с инструкциите на производителя. Накратко 10 uL плазма с 500 uL от 42 тМ H 2 SO 4 се добавя в микро центрофужна епруветка. Добавете 125 μL разтвор на фосфотунгстикова киселина и разбъркайте чрез завихряне. Инкубация при стайна температура за 5 минути, след това центрофугиране за 3 минути при 13 000 × g. Пелетите се събират и се суспендират отново в лед с 100 uL двойно дестилирана 2 О [с 2 мкл ВНТ (100 х)]. Крайният обем се довежда до 200 uL с двойно дестилирана 2 О, и след това 600 мкл от ТВА разтвор във всяка епруветка, съдържаща стандартни и се добавя проба. Инкубация при 95 ° С в продължение на 60 минути. След реакцията с TBA, MDA се определя количествено, като се използва спектрофотометър с дължина на вълната 532 nm [18] .
Концентрацията на глутатион пероксидаза (GSH-PX) в серума беше измерена с помощта на комплект плъхов глутатион пероксидаза Elisa (CUSABIORat GSH-PX ELISA Kit, CSB-E12146r, Университет Wuhan, Ухан, провинция Хубей 430223, КНР). Съгласно инструкциите на производителя беше направена оценка.
МелатонинКонцентрацията на (MT) в серума се изчислява чрез използване на комплект плъхов мелатонин Elisa (CUSABIO MelatoninELISA Kit, CSB-E13433r, Университетска наука Ухан, Ухан, провинция Хубей 430223, КНР). Оценката беше направена съгласно инструкциите на производителя. Както GSH-PX, така и MT измерването се извършват по следната процедура. Накратко бяха приготвени реактиви, проби и стандарт и 100 μl проби и стандарт бяха добавени в ямките и инкубирани 2 часа при 37 ° С. Всяка ямка се отстранява от съдържанието и се добавят 100 μl биотин-антитяло и се инкубира в продължение на 1 час при 37 ° С. Измивайки се три пъти с измиващ разтвор, към всяка ямка се добавя 100 μl HRP-авидин и се инкубира в продължение на 1 час при 37 ° С. След това всяка ямка се промива 5 пъти с промивен разтвор и във всяка ямка се добавя 90 μl TMB субстрат, инкубира се в продължение на 20 минути и накрая към всяка ямка се добавя 50 μl стоп разтвор. Използва се спектрофотометър при дължина на вълната при 450 nm за отчитане на аналитичната плака.
2.7 . Статистически анализ
Всички резултати бяха изразени като средна стойност ± стандартна грешка (средна стойност ± SE). Данните бяха оценени с помощта на статистическата програма SPSS v.19 софтуер (SPSS In. Chicago, IL, USA). Едно- и двупосочен тест ANOVA и LSD е използван за оценка на значимостта между групите и стойностите на P по-малко от 0,05 се считат за значими.
3 . Резултати
3.1 . Телесно тегло на язовира
Тестът ANOVA за телесно тегло на язовирите разкрива, че няма значителни разлики в средното телесно тегло на язовирите в експериментални групи през интервали 1-ва, 2-ра и 3-та седмица.
3.2 . Клинични наблюдения
Клиничният преглед показа, че при някои жени е наблюдавана груба кожа при група от 2 часа / ден на експозиция от интервал от експеримент от 3-та седмица. Аборт и кървене са наблюдавани при една жена от групата на експозиция от 1 h / ден от интервала на експеримента от 3-та седмица. Докато при други жени при експериментални групи с интервали 1-ва и 2-ра седмица не са наблюдавани анормални клинични признаци. Не е отбелязана смърт във всички интервали и групи експерименти. Констатациите след Morton (PM) показват маточна конгестия, кръвоизлив, мъртви и реабсорбирани плодове са наблюдавани и в двете групи на експозиция по време на 2-ри и 3-и интервал от бременността, докато не са наблюдавани констатации на PM при жени от контролните групи ( Фигура 1 ).
3.3 . Фертилитет и пренатално развитие (данни за лапаротомия) за подгрупи A женски
3.3.1 . Продължителност на едноседмична експозиция (GD 0 – GD 7)
Броят на местата за имплантиране и броят на бластоцистите в групата с експозиция от 2 часа на ден са засегнати значително с неравномерно и асиметрично разпределение на местата за имплантиране в двата маточни рога за разлика от контролната група ( Таблица 2 ) ( Фигура 2 ).
Таблица 1 . Ефект на RF-EMR върху телесното тегло на майките през различен период от бременността на експеримента ( Таблица 1 ).
Интервални
групи1-ва седмица 2-ра седмица 3-та седмица Контрол 257.600 ± 8.128 288 700 ± 6,298 323 400 ± 6,172 1 h / ден експозиция 272.200 ± 8.594 275.600 ± 8.466 335.200 ± 3.072 2 h / ден експозиция 257,100 ± 7,004 276.500 ± 11.590 330 700 ± 3,333 P стойност b 0.204 b 0,315 b 0,070 c 0,065 c 0,345 c 0,253 Стойностите са средни ± SE b Сравнение между групата на експозиция 1 h / ден и контролната група, c Сравнение между групата на експозиция 2 h / ден и контролната група, съответно.
Таблица 2 . Ефект на RF-EMR върху пренаталното развитие през различни периоди от бременността.
Групи на параметритеПродължителност на експозицията Една седмица Две седмици Три седмици Брой на местата за имплантиране Брой бластоцисти Брой живи ембриони Брой мъртви ембриони Брой реабсорбирани ембриони Общо не. на плодовете Брой живи плодове Брой на мъртвите плодове Тегло на плода при GD 20 (g) Контрол 10,80 ± 0,89 10,30 ± 0,87 10,20 ± 0,69 0,80 ± 0,32 0,40 ± 0,22 11.30 ± 0.53 10,60 ± 0,61 0,70 ± 0,33 5,670 ± 0,06 1 h / ден експозиция 9,00 ± 0,55 8,40 ± 0,81 7,60 ± 1,36 1,90 ± 0,5 1,30 ± 0,33 7,80 c ± 0,62 7,00 c ± 0,57 0,80 ± 0,24 4,424 c ± 0,09 2 h / ден експозиция 8,50 a ± 0,58 7,90 a ± 0,67 7,20 a ± 0,9 2,30 a ± 0,51 1,60 a ± 0,37 8,30 c ± 0,55 7,50 c ± 0,54 0,80 ± 0,24 4.900 c ± 0.09 P стойност 0,078 0.101 0,085 0.101 0,054 0,001 0,001 0,803 0,001 0,027 0,041 0,049 0,028 0,012 0001 0,001 0,803 0,001 Стойностите са средни ± SE a P <0,05, c P <0,001 в сравнение с контролната група.

Фигура 2 . Интервал 2-ра седмица от бременността. (A) Контролирайте нормалната матка с нормално разпределение на ембрионите в двата маточни рога , (B, C, D) е наблюдавана ранна ембрионална смърт в групите на излагане на RF-EMR и е обозначена със стрелки, (E, H) реабсорбираните ембриони са записани в 1 h / day група експозиция, (F, G) е наблюдавана ранна ембрионална смърт в групите на RF-EMR експозиция, (G, H, J) е наблюдавана умерена до тежка конгестия в маточните рога във всички групи на експозиция. Неравномерното и асиметрично ембрионално разпределение е много ясно при (B, C, D, E, F, G, H и J), за разлика от (A) контрол на маточните рога.
3.3.2 . Двуседмична продължителност на експозицията (GD 0 – GD 14)
Двуседмичното излагане на RF-EMR показва, че средният брой на живите ембриони е значително намален, докато броят на умрелите и реабсорбираните ембриони се увеличава значително в групата на експозиция 2 h / ден в сравнение с контролната група ( Таблица 2 ). Излагането на RF-EMR 1 h / ден няма съществен ефект върху редица живи, мъртви и реабсорбирани ембриони, въпреки че има някои промени в стойностите им. Асиметрично разпределение на ембрионите се наблюдава и в двата маточни рога и в двете групи на експозиция на RF-EMR, с изключение на контролната група ( Фигура 2 ).
3.3.3 . Триседмична продължителност на експозицията (GD 0 – GD 20)
Маточните рога бяха засегнати значително и показаха някои патологични признаци като конгестия и кръвоизлив при облъчени жени от двете групи на експозиция в сравнение с контролната група. Вътрематочната експозиция в продължение на три седмици повлия отрицателно на пренаталното развитие на плода, представено от малформация, хематом в различни части на тялото, увеличаване на дебелината на кожата и едематозни плодове ( Фигура 3 ). Вътрематочното облъчване от 1 800 MHz GSM-подобни сигнали в групи от 1 h / ден и 2 h / ден показва силно значимо намаляване на средния общ брой на плодовете и броя на живите плодове в сравнение с неизлъчените язовири в контролна група, докато редица мъртви плодове остават последователни. Телесната маса на плодовете при GD20 е засегната значително и в двете облъчени групи ( Таблица 2 ).

Фигура 3 . Интервал 3-та седмица от бременността. (A) Контролирайте нормалната матка с нормални плодове, (B, C) 1 час / ден и 2 часа / ден изложената матка на плъхове показва кръвоизлив и задръствания, посочени със стрелка. (D, E, F) GD 20 фетуси показват хематом, мъртви и едематозни плодове при плъхове, изложени на 1 час / ден. (G, H, I) Северна хематома с големи едематозни плодове са наблюдавани при GD 20 на бременността при група с експозиция 2 h / day 2 h / day.
4 . Постнатално развитие (данни за раждането) за бременни жени от подгрупа В
Излагането на RF-EMR вътреутробно през различни интервали от бременността води до различни опасни ефекти върху живородените и тези ефекти включват хематом, груба кожа , зачервяване на кожата , задръствания, къса опашка и малформации и проявите на тези признаци зависят от продължителността на вътрематочната експозиция ( Фигура 4 ).

Фигура 4 . Вътрематочното излагане на RF-EMR показва различни опасни ефекти. (А) Хематом на главата с деформация на главата се наблюдава при кученце след излагане на 1 час / ден в продължение на три седмици. (B) Къса опашка, деформация на главата и хематом на левия крак и опашка се наблюдават при експозиция 1 h / ден в продължение на три седмици. (C) хематом на левия крак при кученце след излагане на облъчване от 1 h / ден. (D, E) хематом в основата на опашката, на долната челюст и зачервяване на кожата с хематом на дясната ръка след 2 h / ден вътрематочно облъчване в продължение на три седмици. (F) претоварени малки с хематом на гърба и основата на опашката на кученце в групата с експозиция 2 h / ден.
4.1 . Данни за доставка
Средният общ брой на новородените малки, броят на живите и мъртвите малки в допълнение към броя на малките с аномалии след вътреутробно облъчване с 1 800 MHz GSM-подобен RF-EMR през първата седмица на бременността са сходни във всичките три групи . След двуседмично вътрематочно излагане данните за раждането показват намаляване на общия брой на малките и броя на живите малки, за разлика от малките от контролната група. Докато мъртвите и ненормалните малки са подобни на контролните. Общият брой на малките и живите малки в групи на експозиция след три седмици на експозиция са значително намалени в сравнение с контролната група и броят на умрелите и нормалните малки е статистически нормален ( Таблица 3 , Таблица 4 , Таблица 5 ).
Таблица 3 . Ефект от едноседмично вътрематочно излагане на RF-EMR върху извънпролетно изследване.
Групи Общо не. на малките Брой живи малки Брой мъртви малки Брой аномалии Контрол 10,20 ± 0,55 10,10 ± 0,54 0,10 ± 0,10 0,30 ± 0,15 1 h / ден експозиция 9,70 ± 0,59 9,50 ± 0,58 0,20 ± 0,13 0,30 ± 0,21 2 h / ден експозиция 9,60 ± 0,61 9,50 ± 0,58 0,10 ± 0,10 0,20 ± 0,13 P стойност b 0,554 b 0,464 b 0,534 б 1.000 c 0,479 c 0,464 c 1000 c 0,681 Стойностите са средни ± SE b Сравнение между групата на експозиция 1 h / ден и контролната група, c Сравнение между групата на експозиция 2 h / ден и контролната група.
Таблица 4 . Ефект от двуседмично вътрематочно излагане на RF-EMR при извънпролетно изследване.
Групи Общо не. на малките Брой живи малки Брой мъртви малки Брой аномалии Контрол 10,50 ± 0,58 10,40 ± 0,60 0,10 ± 0,10 0,10 ± 0,10 1 h / ден експозиция 8,30 ± 0,59 7,90 ± 0,52 0,40 ± 0,22 0,40 ± 0,22 2 h / ден експозиция 6,40 ± 0,60 6,00 ± 0,57 0,40 ± 0,22 0,50 ± 0,16 P стойност b 0,014 b 0,004 b 0,273 b 0,223 c 0,001 c 0,001 c 0,273 c 0,108 Стойностите са средни ± SE b Сравнение между групата на експозиция 1 h / ден и контролната група, c Сравнение между групата на експозиция 2 h / ден и контролната група.
Таблица 5 . Ефект от триседмична RF-EMR вътрематочна експозиция при извънпролетно изследване.
Параметри Общо не. на малките Брой живи малки Брой мъртви малки Брой аномалии Контрол 11.30 ± 0.49 11,10 ± 0,48 0,20 ± 0,13 0,20 ± 0,13 1 h / ден експозиция 7,80 ± 0,74 7,50 ± 0,67 0,30 ± 0,15 0,60 ± 0,22 2 h / ден експозиция 7,50 ± 0,68 7,20 ± 0,64 0,30 ± 0,15 0,50 ± 0,22 P стойност b 0,001 b 0,001 b 0.633 b 0,163 c 0,001 c 0,001 c 0,633 c 0,292 Стойностите са средни ± SE b Сравнение между групата на експозиция 1 h / ден и контролната група, c Сравнение между групата на експозиция 2 h / ден и контролната група.
4.2 . Кученца седмично наддаване на телесно тегло
Средно телесно тегло се взема за 30 малки / група / седмично и се измерва седмично, започвайки от 24 часа след нормалното раждане до 28-дневна възраст (24 часа, D7, D14, D21 и D28). Телесното тегло на кученцето след вътрематочно излагане на RF-EMR през първата седмица от бременността не показва ефект и е сходно във всички експериментални групи ( Фигура 5 ).

Фигура 5 . Ефект на RF-EMR вътрематочна експозиция за една седмица от бременността върху седмичното наддаване на телесно тегло на малките.
Телесното тегло на малките на възраст 24 часа от вътреутробно облъчени язовири в продължение на две седмици не показва значителна разлика в сравнение с контролните малки ( Р = 0,551, Р = 0,335 1 час / ден и 2 часа / ден експозиция съответно). Докато седмичното нарастване на телесното тегло от D 7 – D 28 е значително по-ниско от контролните групи ( P = 0,001). Силно значимо намаление на седмичното наддаване на телесно тегло при новородени малки до D 28 ( P = 0,001) се наблюдава при малки от облъчени вътреутробно майки в продължение на 20 дни и в двете изложени групи в сравнение с контролните малки ( Фигура 6 , Фигура 7 ) .

Фигура 6 . Ефект на RF-EMR вътрематочна експозиция за две седмици от бременността върху седмичното наддаване на телесно тегло на малките.

Фигура 7 . Ефект на RF-EMR вътрематочна експозиция за 20 дни от бременността върху седмичното наддаване на телесно тегло на малките.
4.3 . Нормално физиологично развитие
Нормалното физиологично развитие след раждането се проверява внимателно и индивидуално за 30 малки / група / интервал за проследяване на външния вид на ухото, косата, зъбите, отваряне на очите и определяне на деня на отбиване. Няма забележителни разлики между експерименталните групи и контролната група за постнаталното физиологично развитие след вътрематочна експозиция през първата седмица на бременността ( Фигура 8 ). Не се наблюдава разлика в развитието на външния вид на ухото след двуседмично вътреутробно облъчване, 2 часа / ден вътреутробно облъчване външен вид на косата, развитие на отваряне на очите и ден на отбиване отне повече време от контролните малки, докато след 1 час / ден облъчване, постнаталното физиологично развитие остава последователно ( Фигура 9 ). Три седмици вътреутробноефект на облъчване върху постнаталното физиологично развитие отрицателно чрез увеличаване на дните, необходими за нормален външен вид на косата и предизвикано забавяне на никненето на зъбите и в двете облъчени групи. Отварянето на очите отне по-дълго време, за да се отвори нормално, отколкото контролната група след групата на експозиция 2 h / ден и периодът на отбиване показа увеличение на продължителността на периода на отбиване и в двете вътреутробни групи на облъчване в сравнение с контролната група ( Фигура 10 ).

Фигура 8 . Ефект от едноседмична RF-EMR вътрематочна експозиция върху постнаталното физиологично развитие.

Фигура 9 . Ефект от двуседмична RF-EMR вътрематочна експозиция върху постнаталното физиологично развитие.

Фигура 10 . Ефект от триседмична RF-EMR вътрематочна експозиция върху постнаталното физиологично развитие.
4.4 . Ефект на RF-EMR 1800 MHz подобна на GSM честота върху окислителния и антиоксидантния статус при плъхове през различни периоди на бременност
В това проучване беше изследвано дали 1 800 MHz GSM-подобна честотна експозиция, предизвикана от оксидативен стрес, променя антиоксидантния ензим и хормоналната активност за отстраняване на свободните радикали. Антиоксидантните активности се определят биохимично и с помощта на търговски ELISA комплекти. GD 0 – GD 7 периоди на вътрематочно излагане на RF-EMR показват, че окислителят и антиоксидантната активност във всички експериментални групи остават постоянни. След 2 h / ден вътрематочната експозиция от GD 0-GD 14 GSH-PX активност е значително по-ниска от наблюдението в контролната група. Докато експозицията от 1 h / ден не показва ефект. Нивото на MDA е по-високо значително и в двете групи на експозиция в сравнение с контролната група. Интервалите GD 0 – GD 20 показват значително намаляване на нивото на MT след 2 часа / ден вътрематочна експозиция и значително намаляване на GSH-PX активността и в двете групи на експозиция в сравнение с контролната група. И двете групи за вътрематочно излагане показват значително увеличение на нивото на MDA в сравнение с контролната група. Резултатите са показани в ( Таблица 6).
Таблица 6 . Ефект на 1 800 MHz GSM-подобен RF-EMF върху нивото на мелатонин , активността GSH-PX и нивото на MDA през различния период на бременността. Ефект на 1 800 MHz GSM-подобен RF-EMF върху нивото на мелатонин, активността GSH-PX и нивото на MDA през различния период на бременността.
Групи GD0 – GD7 GD0 – GD14 GD0 – GD20 MT (pg / ml) GSH-PX (mIU / mL) MDA (nmol / ml) MT (pg / ml) GSH-PX (mIU / mL) MDA (nmol / ml) MT (pg / ml) GSH-PX (mIU / mL) MDA (nmol / ml) Контрол 42,966 ± 1,50 64,869 ± 3,61 1,561 ± 0,12 42,221 ± 0,04 74,512 ± 4,71 1,515 ± 0,11 41,0132 ± 1,79 71,253 ± 2,34 1,523 ± 0,09 1 ч / ден Излагане 42,212 ± 1,16 58,614 ± 2,45 1,627 ± 0,17 40,512 ± 0,79 67.411 ± 0.99 1,899 a ± 0,11 38,561 ± 0,48 66.563 a ± 0.99 1,973 a ± 0,17 2 ч / ден Експозиция 40,446 ± 0,77 57,569 ± 0,77 1,823 ± 0,06 40,108 ± 0,90 64,511 a ± 1,34 1,901 a ± 0,12 37.447 a ± 0.81 63,518 c ± 0,84 2.093 c ± 0.10 P стойност 0,657 0,096 0,723 0.201 0,093 0,028 0,133 0,042 0,021 0,145 0,054 0,292 0,117 0,021 0,027 0,035 0,002 0,045 Стойностите са средни ± SE a P < 0,05, c P <0,005 са приети за статистически значими, р стойността е сравнение между контролната група и съответно две третирани групи.
5 . Дискусия
Растежът на плода е един от най-чувствителните етапи на растеж към електромагнитното излъчване, тъй като развиващите се тъкани и органогенезата са по-чувствителни към вредните агенти от тези на възрастните. Досега няма последователни резултати за ефекта на RF-EMR върху пренаталното и постнаталното развитие поради разликата в експерименталния дизайн, включително SAR, честотите, продължителността на експозицията и краткосрочната или дългосрочната експозиция, въпреки че всички честоти, използвани в микровълновата гама. Нашето проучване се различава от предишни проучвания по това, че изследвахме биологичните ефекти на честота 1 800 MHz GSM RF-EMR през различни периоди на бременността и неговия ефект върху растежа и развитиетона ембриони по време на бременност и след бременност. Изяснихме, че 1 800 MHz GSM облъчване по време на бременност е причинило някои вредни ефекти върху пренаталното развитие, представено от значително неравномерно и асиметрично разпределение на местата за имплантиране в двата маточни рога на изложени групи, с изключение на контролната група, намаляване на теглото на плода с някои аномалии, както и индукция на оксидативен стрес при бременни животни, намаляване на седмичното наддаване на телесно тегло и известен отрицателен ефект върху функционалното и физиологичното развитие. Предишни проучвания са разглеждали различни аспекти на въздействието на честота от 1 800 MHz по време на бременност. Например Tomruk et al.установи, че излагането на зайци в средната фаза на бременността в продължение на 15 минути на ден в продължение на седем дни на 1 800 MHz GSM-подобни сигнали води до окислително разрушаване в чернодробната тъкан в резултат на производството на свободни радикали при бременни животни [19] . Друго проучване, проведено от друг турски екип, показва, че експозицията на 1 800 MHz GSM-подобен сигнал на небременни и бременни зайци в продължение на седем дни (15 минути / ден) води до производство на свободни радикали, които предизвикват липидна пероксидация и окислително увреждане на ДНК [15] , Излагане на плода на 800–1 Радиочестотното излъчване с клетъчни телефони с оценка 900 MHz води до промени в поведенческите и неврофизиологичните параметри (нарушена памет, хиперактивност и намалена тревожност), които продължават да съществуват и в зряла възраст, когато мишките са изложени по време на бременност [9] . Нашите констатации бяха подобни на други скорошни проучвания върху ефекта (ите) на експозиция в матката на Wi-Fi честота 2,45 GHz върху бременни животни и техните бебета (преди и след раждането). В едно от такова проучване мишки, изложени на 2,45 GHz EMF в продължение на 2 часа / ден в продължение на 30 дни, тъй като (период на чифтосване 22 дни, период на чифтосване 5 дни и след чифтосване 3 дни) показват асиметрични и неравномерни места за имплантиране в маточните рога в експонирани групи в сравнение с контрола и индуциран пробив на ДНК в мозъчните клетки [20]. В друго проучване мишки, изложени на непрекъсната вълна от 2,45 GHz вътреутробно облъчване 2 h / ден в продължение на 45 дни при SAR 0,023 023 W / kg като (период на чифтосване 20 дни, 5 дни по време на чифтосване и 20 дни след чифтосване) , това проучване показа, че облъчването с MW индуцира оксидативен стрес, който потиска имплантацията (наблюдава се неравномерно / асиметрично разпределение на ембрионите в маточните рога) и води до ембрионална деформация в случай, че бременността продължи и доведе до счупване на веригата на ДНК в мозъчните клетки [21] . Също така нашите констатации са в съгласие с [4], който установи, че бременни плъхове, изложени на 2 450 MHz EMF, водят до ограничаване на постнаталния растеж и забавяне на пубертета при женски плъхове поради индуциране на оксидативен стрес в мозъчните и яйчниковите тъкани. Нашите резултати могат да бъдат интерпретирани, че податливостта към субфертилитет и увреждане на пренаталния растеж поради ранна ембрионална смърт и резорбция на ембриона през втората седмица на бременността е в резултат на вътрематочно облъчване, предизвикващо оксидативен стрес, който играе важна роля в имплантацията и ембрионалния растеж чрез инхибиране на специфични протеини на топлинен шок (Hsp) като Hsp70 и Hsp105. Hsp70 играе критична роля за оплождането и ранното ембрионално развитие при бозайниците. Освен това, Hsp може също да служи като защитна роля в развитието на ембриона, тъй като инхибирането на Hsp70 е причинило намаляване на бластоцистатаразвитие, което може да бъде медиирано от висока степен на апоптоза [22], докато временните и пространствени промени в експресията на Hsp105 в матката на бременни плъхове играят важна физиологична роля в регулирането на имплантацията на ембрион [23]. Констатацията, че RF-EMR влияе на пренаталното развитие, отразява влиянието на много Hsps, трябва да бъде допълнително проучена. Оксидативният стрес се причинява от прекомерното производство на свободни радикали или от дисбаланса на оксидантната / антиоксидантната система. Данните от нашите резултати ясно показват, че RF-EMR (1800 MHz) при ниско ниво на SAR води до намаляване на GSH-PX активността и нивата на MT като антиоксидант ензим и чистач на свободни радикали, както и повишаване на нивото на MDA, биомаркер за липидна пероксидация и тъкан щети поради екстензивно производство на пероксиди и свободни радикали, които причиняват токсичен ефект и увреждат всички клетъчни компоненти, например протеини, липиди и ДНК.
Реактивните кислородни видове (ROS) играят важна роля във физиологичните репродуктивни функции като съзряване, стероидогенеза на яйчниците , функция на жълтото тяло, оплождане, развитие на ембриона и бременност [24] . Въпреки огромните доклади, сочещи, нейонизиращо лъчениеиндуциран оксидативен стрес и неговият ефект върху имплантацията, пренаталното развитие и бременността, прецизният механизъм на действие на RF-EMR при предизвикване на ранна загуба на ембриони, малформации и увреждане на бременността все още е неясен. Трябва да се проведат по-подробни проучвания за определяне на специфичния принос на свободните радикали и Hsps за пренаталното развитие в различните етапи на ембриогенезата. Резултатите от постнаталното развитие показват намаляване на седмичното наддаване на телесно тегло и забавяне на някои функционални и физиологични развития като външен вид на зъбите и косата, отваряне на очите и продължителен период на отбиване. Това може да е резултат от намеса между въздействието на оксидативен стрес и растежни фактори като инсулинови растежни фактори-I IGF-I в организма. Нашият резултат е в съответствие с този на Dundar et al.което показа експозиция от 50 Hz доведе до ограничаване на растежа, забавяне на пубертета и намалено ниво на IGF-I при женски плъхове и вероятно свързано с директен токсичен ефект на електрическото поле върху прицелните органи [25] . IGF-Имам много важна роля във феталното и постнаталното развитие. Разликата в нивата на IGF-I, свързана с експозицията на ЕМП, е докладвана в няколко проучвания. Sangun и сътр. установи, че женските плъхове, изложени на 2,45 GHz по време на бременност, и постнаталното развитие показват намаляване на нивото на IGF-I в постнаталната група в сравнение с контролните животни в резултат на индукция на оксидативен стрес [4] . Picinato и сътр. установи, че МТ чрез активиране на МТ 1 рецептор регулира растежа и диференциацията на панкреатичните островчетачрез активиране на сигналния път на IGF-I и инсулиновите рецептори [26] . През 2009 г. Oner проведе проучване, за да изследва дали MT има роля в превенцията на индуцирана от карбонтетрахлорид хепатотоксичност при плъхове чрез експресия на IGF. Те откриха, че МТ, секретиран от епифизната жлеза, увеличава освобождаването на IGF-I и предотвратява увреждането на черния дроб [27]. Ограничаването на постнаталния растеж в нашия експеримент може да се тълкува като резултат от намаляване на нивото на МТ в експеримента на триседмично излагане на облъчване, което има ефект върху освобождаването на IGF-I, които имат важна роля в постнаталния растеж. Нашите резултати са в контраст с други скорошни проучвания. Например, мишките са били вътреутробно изложени 2 часа / ден в продължение на 14 дни, започвайки след пет дни чифтосване на Wi-Fi сигнал при 2,45 GHz, SAR 4 W / kg и резултатът не е показал ефект върху резултата от бременността поради пренатално излагане на Wi -Fi сигнали [28]. В друго проучване плъховете са били вътреутробно изложени на 2,45 GHz Wi-Fi сигнали (2 часа / ден в продължение на 18 дни при SAR 0,08, 0,4 и 4 W / kg), за да се изследва пре- и постнаталния ефект на Wi-Fi сигналите и не са забелязани аномалии при бременни плъхове, няма признаци на токсичност в пред- и постнаталното развитие на малките [13] . В проучване, проведено във Франция, плъховете са били изложени на 2,45 GHz Wi-Fi сигнали като експозиция на цялото тяло на свободно движещи се плъхове в продължение на 1 час / ден в продължение на 3 седмици (мъжки) две седмици (женски) по време на половото съзряване и изследването предполага че Wi-Fi сигналите нямат ефект върху плодовитостта при мъжете и жените при „стойности на WB SAR до критичното ниво на ICNIRP“ [29] . Настоящото проучване, излагане на RF-EMR 1 800 MHz по време на различни периоди на бременност доведоха до различни ефекти върху резултата от бременността и се забавиха при нормално, функционално и физиологично постнатално развитие при малките от вътрематочни облъчени язовири. Честотата и SAR, използвани в нашия експеримент, бяха в границите на безопасността от указанията на ICNIRP и IEEE. Според резултатите от експеримента, ние предполагаме, че дългосрочното излагане на ЕМП в рамките на честотата на микровълновата печка по време на бременност води до хроничен стрес, който от своя страна може да причини някои вредни ефекти върху пред- и постнаталното развитие по различни пътища, за това трябва да се направят допълнителни изследвания проведени за изясняване на действителните пътища, които включват такива вредни ефекти.
Изявление за конфликт на интереси
Декларираме, че нямаме конфликт на интереси.
Признание
Авторите са длъжни на нашата помощ за научни изследвания в хистопатологична лаборатория за тяхната помощ. Проектът беше изцяло подкрепен от Факултет по ветеринарна медицина на Университет Малайзия Келантан .
Препратки
- [1]
- ICNIIRPУказания за ограничаване на експозицията на вариращи във времето електрически, магнитни и електромагнитни полета (до 300 GHz)Health Phys , 74 ( 4 ) ( 1998 ) , стр. 494 - 522
- [2]
- B. Dasdag , S. Akdag , MZ Kizil , G. Kizil , DU Cakir , B. YokusЕфект на 900 MHz радиочестотно излъчване върху бета амилоиден протеин, протеин карбонил и малондиалдехид в мозъкаElectromagn Biol Med , 31 ( 1 ) ( 2012 ) , стр. 67 - 74
- [3]
- С. Дасдаг , М. ТаЕфект от дългосрочното излагане на радиочестотна радиация от 2,4 GHz, излъчвана от Wi-Fi оборудване, върху функциите на тестиситеElectromagn Biol Med , 34 ( 1 ) ( 2015 ) , стр. 37 - 42
- [4]
- O. Sangun , B. Dundar , H. Darici , S. Comlekci , DK Doguc , S. CelikЕфектите от дългосрочното излагане на 2450 MHz електромагнитно поле върху растежа и развитието на пубертета при женски плъхове WistarElectromagn Biol Med , 8378 ( 2014 ) , стр. 1 - 9
- [5]
- О. Tsybulin , Е. Sidorik , О. Brieieva , L. Buchynska , S. Kyrylenko , D. Henshel , и др.GSM 900 MHz клетъчно излъчване може да стимулира или потисне ранната ембриогенеза при японските пъдпъдъци в зависимост от продължителността на експозициятаInt J Radiat Biol , 89 ( 9 ) ( 2013 ) , стр. 756 - 763
- [6]
- Световна здравна организацияСлужба за преса и връзки с обществеността. Програма за изследване на СЗО за радиочестотни областиСветовна здравна организация , Женева ( 2010 )
- [7]
- JC LinНов IEEE стандарт за нива на безопасност по отношение на излагането на човека на радиочестотно лъчениеIEEE Антени Propag Mag , 48 ( 1 ) ( 2006 ) , стр. 157 - 159
- [8]
- Девентер В. Емили , Е. ван Ронген , Р. СондърсПрограма за изследване на СЗО за радиочестотни областиБиоелектромагнетика , 5 ( 2011 ) , стр. 417 - 421
- [9]
- TS Aldad , G. Gan , XB Gao , HS TaylorИзлагането на радиочестотна радиация на плода от клетъчни телефони с рейтинг 800–1900 MHz влияе върху невроразвитието и поведението на мишкиSci Rep , 2 ( 2012 ) , стр. 312
- [10]
- FAS Ал-КудсиЕфект на електромагнитното подвижно излъчване върху развитието на ембриона на пилетаLife Sci J , 9 ( 2 ) ( 2012 ) , стр. 983 - 991
- [11]
- GG Budak , NB Muluk , GG Öztürk , Б. Budak , A. Апан , N. Seyhan , и др.Ефекти на GSM-подобна радиочестота върху изкривяването на отоакустичните емисии на продукти при бременни възрастни зайциClin Investig Med , 32 ( 2 ) ( 2009 ) , стр. 112 - 116
- [12]
- MJ Yang , JY Liu , YF Wang , HY Lang , X. Miao , LY Zhang , et al.Ефекти на електромагнитния импулс върху полидактилия на миши плодовеТериогенология , 80 ( 1 ) ( 2013 ) , стр. 18 - 23
- [13]
- F. Poulletier de Gannes , E. Haro , A. Hurtier , M. Taxile , A. Athane , S. Ait-Aissa , et al.Ефект на вътреутробната Wi-Fi експозиция върху пред- и постнаталното развитие на плъховеВродени дефекти Res Part B Dev Reprod Toxicol , 95 ( 2012 ) , стр. 130 - 136
- [14]
- D. Средства , KW ChanОценка на спазването на указанията на FCC за излагане на човека на радиочестотни електромагнитни полета: допълнителна информация за оценка на съответствието на мобилните и преносими устройства с ограниченията на FCC за излагане на хората на радиочестотни емисииОфис на инженерната и технологична федерална комисия за комуникации , Вашингтон ( 2001 )
- [15]
- G. Güler , A. Tomruk , E. Ozgur , D. Sahin , A. Sepici , N. Altan , et al.Ефектът на радиочестотното облъчване върху увреждането на ДНК и липидите при женски и мъжки новородени зайциInt J Radiat Biol , 88 ( 4 ) ( 2012 ) , стр. 367 - 373
- [16]
- G. Гюлер , A. Tomruk , Е. Йозгюр , N. SeyhanЕфектът на радиочестотната радиация върху увреждането на ДНК и липидите при небременни и бременни зайци и техните новородениGen Physiol Biophys , 29 ( 2010 ) , стр. 59 - 66
- [17]
- G. Kismali , Е. Йозгюр , G. Гюлер , A. Akcay , Т. Sel , N. SeyhanВлиянието на 1800 MHz GSM-подобни сигнали върху кръвната химия и оксидативния стрес при небременни и бременни зайциInt J Radiat Biol , 88 ( 2012 ) , стр. 414 - 419
- [18]
- NA Botsoglou , DJ Fletouris , GE Papageorgiou , VN Vassilopoulos , AJ Mantis , AG TrakatellisБързо чувствителен и специфичен метод на тиобарбитурова киселина за измерване на липидната пероксидация в животински тъкани, храни и фуражни пробиJ Agric Food Chem , 42 ( 1994 ) , стр. 1931 - 1937
- [19]
- А. Томрук , Г. Гюлер , А. С. ДинселВлиянието на 1800 MHz GSM-подобни сигнали върху чернодробната окислителна ДНК и увреждане на липидите при небременни, бременни и новородени зайциCell Biochem Biophys , 56 ( 2010 ) , стр. 39 - 47
- [20]
- VP Singh , P. Singh , CM Chaturvedi , RK Shukla , A. Dhawan , RK Gangwar , et al.2,45 GHz ниско ниво на CW микровълнова радиация засяга местата за имплантиране на ембриони и едноверижно увреждане на ДНК в мозъчните клетки на мишки, mus musculus2009 Международна конференция за нововъзникващите тенденции в електронните и фотонните устройства и системи , IEEE , Варанаси ( 2009 ) , стр. 379 - 382
- [21]
- S. Shahin , VP Singh , RK Shukla , A. Dhawan , RK Gangwar , SP Singh2.45 GHz индуциран от микровълновото облъчване оксидативен стрес засяга имплантирането или бременността при мишки, mus musculusAppl Biochem Biotechnol , 169 ( 5 ) ( 2013 ) , стр. 1727 - 1751
- [22]
- Matwee WAK. Кристи , Мусадин Камарудин , Дийн Х. Бетс , ПК БасрЕфектите на антителата към протеина на топлинен шок 70 при оплождането и развитието на ембрионаMol Hum Reprod , 7 ( 9 ) ( 2001 ) , стр. 829 - 837
- [23]
- JX Yuan , LJ Xiao , CL Zhang , XS Lu , T. Liu , M. Chen , et al.Повишена експресия на протеин от топлинен шок 105 в матката на плъх в ранна бременност и неговото значение при имплантиране на ембриониReprod Biol Endocrinol , 7 ( 23 ) ( 2009 ) , стр. 1 - 10
- [24]
- А. Агарвал , С. Гупта , РК ШармаРоля на оксидативния стрес в женската репродукцияReprod Biol Endocrinol , 21 ( 2005 ) , стр. 1 - 21
- [25]
- Б. Dundar , G. Cesur , S. Comlekci , A. Songur , A. Gokcimen , О. Sahin , и др.Ефектът на пренаталното и постнаталното дългосрочно излагане на електрическо поле от 50 Hz върху растежа, развитието на пубертета и нивата на IGF-1 при женски плъхове WistarToxicol Ind Health , 25 ( 7 ) ( 2009 ) , стр. 479 - 487
- [26]
- MC Picinato , AE Hirata , J. Cipolla-Neto , R. Curi , CRO Carvalho , GF Anhê , et al.Активиране на инсулинови и IGF-1 сигнални пътища от мелатонин чрез MT1 рецептор в изолирани острови на панкреаса на плъхJ Pineal Res , 44 ( 1 ) ( 2008 ) , стр. 88 - 94
- [27]
- Жале Oner , Илтер Кус , Хакан OnerМелатонинът увеличава експресията на инсулиноподобен растежен фактор I при плъхове с индуцирано от карбонтетрахлорид чернодробно уврежданеJ Anim Veterin Adv , 8 ( 11 ) ( 2009 ) , стр. 2256 - 2261
- [28]
- М. Sambucci , F. Laudisi , F. Nasta , R. Pinto , R. Lodato , P. Altavista , и др.Пренатално излагане на нейонизиращо лъчение: ефекти на wifi сигналите върху резултата от бременността, периферното отделение на b-клетките и производството на антителаRadiat Res , 174 ( 2010 ) , стр. 732 - 740
- [29]
- F. Poulletier de Gannes , B. Billaudel , E. Haro , M. Taxile , L. Le Montagner , A Hurtier , et al.Плодовитост на плъхове и ембрионално развитие на плода: влияние на излагането на Wi-Fi сигналаReprod Toxicol , 36 ( 2013 ) , стр. 1 - 5
Използване на мобилен телефон по време на бременност и риск от спонтанен аборт. Mahmoudabadi, Ziaei, et al. Вестник за екологично здраве и наука и инженерство 13:34 (2015).
Използване на мобилен телефон по време на бременност и риск от спонтанен аборт
Въведение
Повдигнати са широко разпространени опасения относно излагането на електромагнитно поле (ЕМП) от източници, използвани за мобилни телекомуникации [ 1 - 4]. Голяма част от безпокойството възниква, защото новите технологии се въвеждат без адекватно предоставяне на публична информация за тяхното естество или обсъждане на дебата в научната общност относно възможните последици за здравето. Всъщност използването на мобилни телефони се е увеличило значително заедно с намаляването на разходите му и развиващите се страни създават мобилни телекомуникации, а не по-скъпите фиксирани системи. По този начин, ако има въздействие върху здравето от мобилните телефони, това ще засегне всички по света. Основният фокус на изследванията е върху радиочестотното (RF) лъчение, което се генерира главно от телефони, докато някои учени са загрижени за възможното въздействие на излагането на полета с изключително нискочестотно електромагнитно поле (ELF-EMF), генерирани от захранващи токове в телефона [ 5- 7 ]. Устройството с най-голяма консумация на енергия е предният усилвател. Следователно, съответният ELF-EMF има спектър, подобен на импулсната структура на RF сигналите.
Общественото обсъждане на опасностите за здравето от излагане на ЕМП се фокусира върху възможната връзка с рак, сърдечно-съдова и имунна система. По-малко внимание е отделено на оценката на оценката на рисковете за здравето от експозиция на ЕМП върху репродуктивното здраве, като спонтанни аборти [ 7 - 9 ].
Здравният ефект на ЕМП върху бременността остава спорен, въпреки усилията за постигане на консенсус [ 10 ]. Основните предизвикателства при изучаването на ЕМП са; 1) точна оценка през съответния период от време и; 2) идентифициране на податлива популация.
Поради това е разработено проучване за контрол на случая, за да се определят ефектите от излагането на ЕМП на мобилни телефони веднага след спонтанни аборти при избрани пациенти.
Методи
За оценка на ефектите от излагането на ЕМП на мобилни телефони върху спонтанните аборти беше избрано проучване за контрол на случая. Поради липсата на знания за тези ефекти е проведено пилотно проучване върху 100 участници. След това, въз основа на първичните данни, с ниво на значимост α = 0,05 и мощност 1-β = 0,80, беше изчислен размер на извадката от 220 участници за всяка група. В настоящото проучване на случай-контрол са включени 292 жени, които са имали необясним спонтанен аборт при <14 гестационна седмица (като ранен аборт) непосредствено след аборт (през първите 48 часа) и 308 бременни жени при 14 гестационна седмица. Гестационната възраст се изчислява въз основа на последния менструален цикъл, потвърден от ултразвуково изследване. Субектите бяха наети от 10 болници в Техеран.
Ситуациите по време на бременност при бременните жени в контролната група бяха съпоставени с възрастта на майката и бащата, гравитацията, ИТМ преди бременността, семейните отношения, продължителността от последното раждане, образователното ниво, професията и историята на предишни аборти или преждевременно раждане с жените в група дела.
Критериите за включване бяха следните:
едноплодна бременност; 2) 18–35 годишна възраст; 3) спонтанна бременност без използване на асистирана репродуктивна технология
Критериите за изключване бяха следните:
хронични заболявания (диабет, хипертония, заболявания на щитовидната жлеза, заболявания на имунната система и сърдечно-съдови заболявания); 2) всякакви генетични нарушения (при жените, техните съпрузи и техните роднини от първа степен); 3) вагинално кървене през първия триместър на бременността в контролната група; 4) история на вродени дефекти при предишни бременности; и 5) пушене на цигари, алкохол или консумация на наркотици.
Жените попълниха формуляра за събиране на данни, който беше използван за събиране на данни за социално-икономически и акушерски характеристики, медицинска и репродуктивна история и начин на живот. Вторият формуляр за събиране на данни, попълнен от обучен интервюиращ, е предназначен да получи информация за използването на мобилни телефони по време на бременност. За разглеждане на ефектите от мобилните телефони измерихме средното време за разговори на ден, местоположението на мобилните телефони, когато не се използват (в чанти, джобове за дрехи или ≥60-70 см от тялото), използване на ръце- безплатно оборудване, използване на телефони за други приложения (за изпращане на съобщения, слушане на музика и игра на игри), специфичната скорост на усвояване (SAR), отчетена от производителя, и средната стойност на ефективната SAR (средна продължителност на времето за разговор на ден × SAR). SAR показва колко електромагнитна енергия се абсорбира от телесните тъкани. Този параметър показва оценката на общата ЕМП енергия. Ефективният SAR може да покаже косвено излагането на оценка на ELF от мобилни телефони. Проучихме данните за SAR за всеки модел мобилен телефон от сайта SAR.ir.
За прецизна оценка на времето за разговори бяха разгледани разписките от сметките за мобилни телефони. Информацията за времето, прекарано за далекосъобщения в конкретния раздел на новите версии на мобилните телефони, също беше взета под внимание.
Процедурата е одобрена от Комитета по етика на университета Tarbiat Modares. Всички жени участваха доброволно и предоставиха писмено информирано съгласие.
Статистически анализ
Демографските и акушерските характеристики, както и параметрите, които отразяват използването на клетъчни телефони, бяха сравнени между двете групи, използвайки t-тестове и хи-квадрат тестове. Използван е логистичен регресионен модел за оценка на връзката между спонтанните аборти като бинарен зависим фактор и ефективната SAR, възрастта на майката, възрастта на бащата, историята на абортите и семейните отношения като независими фактори. Анализите бяха извършени със статистически софтуер за състояние на пакета (SPSS) v.16. Стойността на P <0,05 се считаше за статистически значима. Всички интервали на доверие са изчислени на ниво 95%.
Резултати
Разпределението на демографските и потенциалните смутители за спонтанни аборти е представено в таблица 1. Както беше показано, няма значителни разлики в възрастта на майката и бащата, ИТМ преди бременността, професията и образователния статус, социално-икономическото ниво, семейните отношения, продължителността от последното раждане и историята на предишни аборти или преждевременни трудове между двете групи.
маса 1
Сравнението на характеристиките на майката и бащата между двете групи
Параметри Група дела (n: 226) Контролни групи (n: 246) P Възраст на майката (години) * Средно ± SD 27,81 ± 5,20 27,34 ± 4,3 0.2 Възраст на бащата (години) * Средно ± SD 32,62 ± 5,46 31,90 ± 5,49 0,1 Семейни отношения ** N (%) 62 (21,24) 77 (25,1) 0,3 ИТМ преди бременността * 25,14 ± 6,98 24,29 ± 5,24 0,09 История на абортите ** N (%) 47 (20,8) 36 (14,6) 0.2 История на преждевременно раждане ** N (%) 3 (1,3) 2 (0,8) 0.6 Продължителност от последната доставка (години) * Средно ± SD 6,61 ± 3,93 6,57 ± 3,46 0.9 Професия N (%) ** 0.2 Домакиня 195 (86,4) 225 (91,5) Служител 31 (13,6) 21 (8,5) Образователен статус N (%) ** 0,09 Начално училище 88 (38,9) 98 (39,8) Средно и средно училище 94 (41,6) 100 (40,6) университет 44 (19,5) 48 (19,6) * T студентски тест ** Тест хи квадрат.
Както е показано в табл 2, всички данни, свързани с използването на мобилни телефони, като средно време за разговори на ден, местоположението на мобилните телефони, когато не се използват, използване на телефони за други приложения, специфичната скорост на усвояване (SAR) и средната стойност на ефективна SAR), с изключение на това, че използването на устройства за свободни ръце не е различно между двете групи (p <0,001). Логистичният регресионен анализ разкри значителна връзка между ефективния SAR с риска от спонтанни аборти след корекция за възрастта на майката, възрастта на бащата, историята на абортите и семейните отношения (ИЛИ: 1.11, p <0.001) (Таблица 3).
Таблица 2
Сравнението на употребата на мобилен телефон по време на бременност между двете групи
Параметри Група дела (n: 226) Контролна група (n: 246) P (2 опашки) Време за разговори на ден * (минути) Средно ± SD 9,31 ± 15,22 3,14 ± 3,59 <0,001 Използване на свободни ръце ** n (%) 19 (8,4) 11 (4,5) 0,09 местоположение на телефони, когато не се използват ** n (%) <0,001 на маси 86 (38,1) 77 (31,3) в чанти 101 (44,7) 153 (62,2) в джоба 39 (17,3) 16 (6,5) използване на телефон за други приложения ** n (%) 90 (39,8) 44 (17,9) <0,001 Ефективна SAR * Средна стойност ± SD 7,02 ± 13,92 2,28 ± 3,20 <0,001 * T тест за студент, ** тест с хи квадрат или точен тест на Фишър, SAR: Специфична степен на абсорбция.
Таблица 3
Асоциация на спонтанните аборти с ефективната SAR (специфична степен на абсорбция) *
ИЛИ (95% ДИ) Б. SE P Ефективна SAR 1,11 (1,07-1,16) 0.106 0,21 <0,001 * Коригирано за възрастта на майката, възрастта на бащата, историята на абортите и роднините на съпруга / съпругата.
Дискусия
Излагането на ЕМП е все по-често, но потенциалното влияние върху здравето не е подробно определено, особено при бременни жени. Електромагнитните полета могат да произвеждат биологичен стрес и свободни радикали, които могат да направят податлива популация податлива на вродени малформации и увреждане на тъкани и клетки [ 11 ]. Дългосрочното излагане на електромагнитни полета може да бъде свързано с още по-високи нива на оксидативен стрес, с гореспоменатите съответни промени.
Има ограничена информация за връзката между експозицията на ЕМП по време на бременност и репродуктивните резултати [ 10 - 18 ]. Някои проучвания съобщават за повишен риск от спонтанни аборти и вродени малформации, въпреки че тези резултати са получени от лошо проектирани проучвания.
Това проучване демонстрира повишен риск от спонтанни аборти, свързани с експозиция на ЕМП, и потвърди резултатите от други изследвания [ 10 , 12 ]. Лий проведе вложено проучване за контрол на случая, за да оцени връзката на жилищните и личните магнитни полета със спонтанни аборти [ 12]. Той съобщава, че максималното излагане на лично магнитно поле е свързано с риска от клинични спонтанни аборти. Имаше едно основно ограничение за това проучване; по-специално, измерванията на магнитното поле са получени месеци след първия триместър и появата на спонтанни аборти. Може би поведението и магнитните полета на жените, които са имали спонтанни аборти, са различни от контролите на бременните. Но нашето проучване имаше тази сила, че данните бяха получени малко след спонтанните аборти в групата на случаите и в началото на второто тримесечие в контролната група. Това доведе до по-добра оценка на условията на околната среда и начина на живот на участниците
Въпреки това, настоящите констатации са в противоречие с резултатите от някои други трудове [ 14 - 18 ]. Някои изследвания не поддържат връзка между електрическото и магнитното поле и репродуктивните увреждания. Много от тях обаче са изследвали ефектите на ЕМП или върху животни, или in vitro, а не върху хора [ 14 - 16 ]. Също така, липсата на доказателства за силна връзка между ЕМП и неблагоприятните репродуктивни резултати може да бъде преди всичко резултат от наличието на малко информационни данни поради липсата на епидемиологични данни в тази област.
Трябва да се има предвид едно потенциално ограничение, когато човек интерпретира резултатите от настоящото проучване. Проучването не е оценило всички неизвестни рискови фактори за спонтанни аборти, като балансирани хромозомни аномалии при един родител. Също така, данните за неизвестен спонтанен аборт на много ранен етап не бяха събрани. Друго ограничение на това проучване е неговият характер на контрола на случая и това може да означава, че трябва да се внимава при причинно-следствена интерпретация на констатациите. Третото ограничение на това проучване е, че мобилните телефони може да не са единственият източник на ЕМП. Тъй като много параметри, които могат да повлияят на експозицията на ЕМП и стилове на живот като социално-икономически статус, са били съпоставени между двете групи, има съмнения, че вероятно ефектите, наблюдавани в това проучване, се дължат на използването на мобилни телефони.
Връзката между използването на мобилни телефони и риска от спонтанни аборти срещу потенциални объркващи фактори беше подкрепена с доказателства, че въпреки корекциите за много известни или предполагаеми рискови фактори в логистичните регресионни анализи, оценката не е променена значително.
Въпреки че механизмите, залегнали в ефектите на ЕМП върху риска от спонтанни аборти, не са добре разбрани, ранните ембриони са известни като чувствителни към експозицията на околната среда. Неблагоприятният ефект по време на ранното развитие на плода на клетъчно ниво от EMF на клетъчните телефони може да доведе до смърт на плода. EMF в мобилните телефони е както ELF-EMF, така и RF-EMF. Въз основа на разстоянието от тялото до вътрешността на матката, експозицията, достигаща плода, вероятно е изключително нискочестотно електромагнитно излъчване.
Заключение
Въпреки липсата на ясно разбиране на основните механизми, настоящият резултат предполага, че използването на мобилни телефони може да е свързано с ранни спонтанни аборти, поради което е необходимо по-нататъшно проучване.
Благодарности
Това проучване е проведено с любезното съдействие на участващите пациенти. Освен това, това проучване е финансирано от университета Tarbiat Modares, Техеран, Иран.
Бележки под линия
Конкуриращи се интереси
Авторите заявяват, че нямат конкуриращи се интереси.
Принос на авторите
FSM извърши проектиране и събиране на данни на изследването, SZ извърши дизайн на изследването и подготви статията, MF участва в дизайна на изследването, AK препоръча статистическия дизайн на изследването. Всички автори прочетоха и одобриха окончателния ръкопис.
Информация за сътрудника
Фатеме Шамси Махмудабади, имейл: ri.ca.seradom@f.ismahs .
Saeideh Ziaei, имейл: ri.ca.seradom@as_ieaiz .
Мохамад Фирузабади, имейл: ri.ca.seradom@rimruop .
Аноширван Каземнежад, имейл: moc.oohay@ililka .
Препратки
1. Repacholi MH. Рискове за здравето от използването на мобилен телефон. Здраве на труда и околната среда. 1211 Женева 27, Швейцария: Световната здравна организация; 2001. [ Google Scholar ]2. Divan HA, Kheifets L, Obel C, Olsen J. Пренатално и постнатално излагане на използването на мобилен телефон и поведенчески проблеми при деца. Епидемиология. 2008; 19 (4): 1–7. doi: 10.1097 / EDE.0b013e318175dd47. [ PubMed ] [ CrossRef ] [ Google Scholar ]3. Toropainen A. Експозиция на хора от мобилни телефони в затворена зона. Биоелектромагнетика. 2003; 24 (10): 63–5. doi: 10.1002 / bem.10087. [ PubMed ] [ CrossRef ] [ Google Scholar ]4. Wilen J, Sandstrom M, Hansson MK. Субективни симптоми сред потребителите на мобилни телефони-Последица от поглъщането на радиочестотното поле? Биоелектромагнетика. 2003; 24 (3): 152–9. doi: 10.1002 / bem.10101. [ PubMed ] [ CrossRef ] [ Google Scholar ]5. Робърт Е. Вътреутробни ефекти на електромагнитните полета (нискочестотни, средночестотни RF и микровълнови): преглед на епидемиологичните проучвания. Тератология. 1999; 59 : 292–8. doi: 10.1002 / (SICI) 1096-9926 (199904) 59: 4 <292 :: AID-TERA14> 3.0.CO; 2-8. [ PubMed ] [ CrossRef ] [ Google Scholar ]6. Bernabo N, Tellamanti E, Russo V, Martelli A, Turriani M, Mattoli M, et al. Излагането на електромагнитно поле с изключително ниска честота влияе върху резултата от оплождането в модела на свине. Териогенология. 2010; 73 (9): 1293–305. doi: 10.1016 / j.theriogenology.2009.12.010. [ PubMed ] [ CrossRef ] [ Google Scholar ]7. Jauchem JR. Ефекти от ниско ниво на радиочестотна енергия (3 KHz до 300 GHz) върху човешката сърдечно-съдова, репродуктивна, имунна и други системи: Преглед на скорошната литература. Int J Hyg Environment Health. 2008; 211 (1–2): 1–29. doi: 10.1016 / j.ijheh.2007.05.001. [ PubMed ] [ CrossRef ] [ Google Scholar ]8. Ердоган Ю. Електромагнитно замърсяване в компютърните лаборатории: Ефектите върху учебната среда. Есета Educ. 2007; 22 : 78–86. [ Google Scholar ]9. Ahlbom A, Cardis E, Green A, Linet M, Savitz D, Swerdlow A. Преглед на епидемиологичната литература за ЕМП и здравето. Околна среда. Здравен Перспект. 2001; 109 (6): 911–33. doi: 10.2307 / 3454653. [ PMC безплатна статия ] [ PubMed ] [ CrossRef ] [ Google Scholar ]10. Li DK, Odouli R, Wi S, Janevic T, Golditch I, Bracken TD, et al. Популационно проспективно кохортно проучване на лична експозиция на магнитни полета по време на бременност и риск от спонтанен аборт. Епидомиология. 2002; 13 : 9–20. doi: 10.1097 / 00001648-200201000-00004. [ PubMed ] [ CrossRef ] [ Google Scholar ]11. Шоу ГМ. Неблагоприятни репродуктивни резултати при човека и електромагнитни полета: кратко резюме на епидемиологичната литература . Биоелектромагнетика. 2001; 5 : S5–18. doi: 10.1002 / 1521-186X (2001) 22: 5 + <:: AID-BEM1020> 3.0.CO; 2-L. [ PubMed ] [ CrossRef ] [ Google Scholar ]12. Lee GM, Neutra RR, Hristova L, Yost M, Hiatt RA. Вложено изследване на случай на контрол на жилищни и лични измервания и спонтанни аборти. Епидемиология. 2002; 13 (1): 21–31. doi: 10.1097 / 00001648-200201000-00005. [ PubMed ] [ CrossRef ] [ Google Scholar ]13. Savits DA. Магнитни полета и спонтанен аборт. Епидемиология. 2002; 13 (1): 1–3. doi: 10.1097 / 00001648-200201000-00001. [ PubMed ] [ CrossRef ] [ Google Scholar ]14. Khaki AA, Khaki A, Garachurlou S, Khorshidi F, Tajadini N, Madinei N. Преди и след раждането на 50 HZ електромагнитни полета върху простатните жлези на плъхове: проучване с електронна микроскопия. IJRM. 2008; 6 (2): 77–82. [ Google Scholar ]15. Sun W, Tan Q, Pan Y, Fu Y, Sun H, Chiang H. Ефекти от 50-Hz експозиция на магнитно поле върху хормонална секреция и свързана с апоптоза генна експресия при човешки първи триместър вилозни трофобласти in vitro. Биоелектромагнетика. 2010; 31 (7): 566–72. doi: 10.1002 / bem.20596. [ PubMed ] [ CrossRef ] [ Google Scholar ]16. Huuskonen H, Saastamoinen V, Komulainen H, Laitinen J, Juutilainen J. Ефекти на нискочестотните магнитни полета върху имплантирането при плъхове. Репродуктивна токсикология; 2000; 15 (1): 49–59 [ PubMed ]17. Marcus M, McChesney R, Golden A, Landrigan P. Терминали за видео дисплей и спонтанен аборт. J Am Med Womens Assoc. 2000; 85 (2): 84–8. [ PubMed ] [ Google Scholar ]18. Parazzini F, Chiaffarino F, Chatenoud L, Cipriani S, Ricci E, Chiantera V, et al. Излагане на терминали за видео дисплей и риск от раждане в малка гестационна възраст. J Environment Health. 2010; 72 (6): 24–7. [ PubMed ] [ Google Scholar ]Окислителни механизми на биологичната активност на радиочестотното лъчение с ниска интензивност. Yakymenko, et al. Електромагнитна биология и медицина 34 (3): 1-16 (2015).
Окислителни механизми на биологичната активност на радиочестотното лъчение с ниска интензивност
Принадлежности- PMID: 26151230
- DOI: 10.3109 / 15368378.2015.1043557
Резюме
Настоящият преглед има за цел да обхване експериментални данни за окислителните ефекти на радиочестотното лъчение с ниска интензивност (RFR) в живите клетки. Анализът на наличната в момента рецензирана научна литература разкрива молекулярни ефекти, индуцирани от RFR с ниска интензивност в живите клетки; това включва значително активиране на ключови пътища, генериращи реактивни кислородни видове (ROS), активиране на пероксидация, окислително увреждане на ДНК и промени в активността на антиоксидантните ензими. Това показва, че сред 100 налични в момента рецензирани проучвания, занимаващи се с окислителни ефекти на RFR с ниска интензивност, като цяло 93 потвърждават, че RFR предизвиква окислителни ефекти в биологичните системи. Широкият патогенен потенциал на индуцираните ROS и тяхното участие в клетъчните сигнални пътища обяснява редица биологични / здравни ефекти от RFR с ниска интензивност, които включват както ракови, така и неракови патологии. В заключение, нашият анализ показва, че RFR с ниска интензивност е експресивен окислител за живи клетки с висок патогенен потенциал и че оксидативният стрес, предизвикан от излагане на RFR, трябва да бъде признат за един от основните механизми на биологичната активност на този вид радиация.
Ключови думи: клетъчна сигнализация; рак; свободни радикали; оксидативен стрес; радиочестотна радиация; реактивни кислородни видове.
Подобни статии
- Преки и косвени ефекти от излагане на 900 MHz GSM радиочестотни електромагнитни полета върху клетъчната линия на СНО: Доказателство за страничен ефект от нейонизиращо лъчение.Environment Res. 2019 юли; 174: 176-187. doi: 10.1016 / j.envres.2019.03.063. Epub 2019 26 април.PMID: 31036329
- Свръхпродукция на видове свободни радикали в ембрионални клетки, изложени на радиочестотна радиация с ниска интензивност.Exp Oncol. 2013 септември; 35 (3): 219-25.PMID: 24084462
- Влиянието на 1800 MHz GSM-подобни сигнали върху химията на кръвта и оксидативния стрес при небременни и бременни зайци.Int J Radiat Biol. 2012 май; 88 (5): 414-9. doi: 10.3109 / 09553002.2012.661517. Epub 2012 2 март.PMID: 22280439
- Ефект на радиочестотната радиация върху репродуктивното здраве.Indian J Med Res. 2018 декември; 148 (Suppl): S92-S99. doi: 10.4103 / ijmr.IJMR_1056_18.PMID: 30964085 Безплатна статия от PMC. Преглед.
- Свободни радикали, метали и антиоксиданти при рак, предизвикан от оксидативен стрес.Chem Biol Interact. 2006 г. 10 март; 160 (1): 1-40. doi: 10.1016 / j.cbi.2005.12.009. Epub 2006 23 януари.PMID: 16430879 Преглед.
Цитирано от 25 статии
- Здравните рискове от радиочестотната радиация, включително 5G, трябва да се оценяват от експерти без конфликт на интереси.Онкол Лет. 2020 г. октомври; 20 (4): 15. doi: 10.3892 / ol.2020.11876. Epub 2020 15 юли.PMID: 32774488 Безплатна статия от PMC.
- Призиви, които имат значение или не, за мораториум върху разполагането на пето поколение, 5G, за микровълнова радиация.Mol Clin Oncol. 2020 март; 12 (3): 247-257. doi: 10.3892 / mco.2020.1984. Epub 2020 22 януари.PMID: 32064102 Безплатна статия от PMC.
- Генетичната възприемчивост може да промени връзката между употребата на мобилен телефон и рака на щитовидната жлеза: Проучване, основаващо се на популация, в КънектикътEnvironment Res. 2020 март; 182: 109013. doi: 10.1016 / j.envres.2019.109013. Epub 2019 6 дек.PMID: 31918310
- Излагането на радиочестота на ниско ниво не предизвиква промени в биологията на MSC: Проучване in vitro за предотвратяване на увреждания, свързани с NIR.Клониране на стволови клетки. 2019 декември 18; 12: 49-59. doi: 10.2147 / SCCAA.S204166. eCollection 2019.PMID: 31908499 Безплатна статия от PMC.
- 5G технология: Защо трябва да очакваме преминаване от индуцирани от RF мозъчни ракове към рак на кожата?J Biomed Phys Eng. 2019 1 октомври; 9 (5): 505-506. doi: 10.31661 / jbpe.v0i0.1225. eCollection 2019 октомвриPMID: 31750263 Безплатна статия от PMC.
Ефекти от експозицията на пренатално 900 MHz електромагнитно поле върху хистологията на бъбреците на плъхове. Ulubay, et al. Международен вестник по радиационна биология 91 (1): 35-41
Ефекти от пренаталната експозиция на електромагнитно поле от 900 MHz върху хистологията на бъбреците на плъхове
Принадлежности- PMID: 25084839
- DOI: 10.3109 / 09553002.2014.950436
Резюме
Цел: Да се изследват вредните ефекти от пренаталната експозиция на 900 мегагерцово (MHz) електромагнитно поле (EMF) върху бъбреците на четириседмични мъжки плъхове и да се определят защитните ефекти на мелатонин (MEL) и омега-3 (ω-3) .
Материали и методи: Двадесет и един плъхове албинос на Wistar бяха разпределени на случаен принцип в седем групи, както следва: Control (Cont), Sham, MEL, ω-3, EMF, EMF + MEL и EMF + ω-3. След чифтосването три групи (EMF, EMF + MEL, EMF + ω-3) бяха изложени на EMF. През четвъртата седмица след раждането, произволно бяха избрани шест плъха от всяка група. Средният обем на бъбреците и бъбречните кори, общият брой гломерули и основната хистологична структура на бъбреците са оценени съответно със стереологични и светлинни микроскопични методи.
Резултати: Стереологичните резултати определят, че средният обем на бъбреците и корите са значително увеличени в групи, изложени на ЕМП в сравнение с групата Cont. Неекспонираните групи с ЕМП обаче не са значително модифицирани в сравнение с групата Cont. Освен това общият брой гломерули е значително по-висок при неекспонирани групи с ЕМП в сравнение с групата Cont. Алтернативно, броят на гломерулите в групи, изложени на ЕМП, е намален в сравнение с групата Cont.
Заключения: Пренаталното излагане на бъбреците на плъхове на 900 MHz EMF води до увеличаване на общия обем на бъбреците и намаляване на броя на гломерулите. Освен това MEL и ω-3 предотвратяват неблагоприятните ефекти на ЕМП върху бъбреците.
Ключови думи: EMF-експозиция; хистопатология; развитие на бъбреците; плъх; стереология.
Подобни статии
- Невропротективни ефекти на мелатонин и омега-3 върху хипокампалните клетки, пренатално изложени на 900 MHz електромагнитни полета.Int J Radiat Biol. 2016 октомври; 92 (10): 590-5. doi: 10.1080 / 09553002.2016.1206223. Epub 2016 21 юли.PMID: 27442260
- Ефекти от пренаталното излагане на 900 MHz електромагнитно поле върху зъбната извивка на плъхове: стереологично и хистопатологично проучване.Brain Res. 2008 г. 31 октомври; 1238: 224-9. doi: 10.1016 / j.brainres.2008.08.013. Epub 2008 16 август.PMID: 18761003
- Има ли омега-3 защитен ефект върху надбъбречната жлеза на плъх, изложена на 900 MHz електромагнитни полета?J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 185-190. doi: 10.1016 / j.jmau.2017.08.003. Epub 2017 21 септември.PMID: 30023253 Безплатна статия от PMC.
- Излагане на ниско ниво на радиочестотни електромагнитни полета: ефекти върху здравето и нужди от изследвания.Биоелектромагнетика. 1998; 19 (1): 1-19.PMID: 9453702 Преглед.
- Клетъчната основа на бъбречно увреждане от лъчение.Br J Рак Доп. 1986; 7: 257-64.PMID: 3521704 Безплатна статия от PMC. Преглед. Няма наличен резюме.
Цитирано от 6 статии
- Радиации и мъжка плодовитост.Reprod Biol Endocrinol. 2018 декември 9; 16 (1): 118. doi: 10.1186 / s12958-018-0431-1.PMID: 30445985 Безплатна статия от PMC. Преглед.
- Ефекти на 900-MHz излъчване на хипокампа и малкия мозък на възрастни плъхове и намаляване на тези ефекти от фолиева киселина и Boswellia Сакра .J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 216-224. doi: 10.1016 / j.jmau.2017.09.003. Epub 2017 18 септември.PMID: 30023257 Безплатна статия от PMC.
- Ефекти на фолиевата киселина върху бъбреците на плъхове, изложени на 900 MHz електромагнитно излъчване.J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 198-205. doi: 10.1016 / j.jmau.2017.06.001. Epub 2017 17 юни.PMID: 30023255 Безплатна статия от PMC.
- Ефекти от излагането на електромагнитни полета върху антиоксидантната защитна система.J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 167-176. doi: 10.1016 / j.jmau.2017.07.003. Epub 2017 2 август.PMID: 30023251 Безплатна статия от PMC. Преглед.
- Грижи преди зачеването: нов стандарт на грижи в рамките на здравните услуги за майките.Biomed Res Int. 2016; 2016: 6150976. doi: 10.1155 / 2016/6150976. Epub 2016 29 май.PMID: 27314031 Безплатна статия от PMC.
(2015).
Ефектът от експозицията на плъхове по време на пренаталния период на радиационно разпространение от мобилни телефони върху бъбречното развитие. Bedir, et al. Бъбречна недостатъчност 37 (2): 305-9 (2014).
Ефектът от излагането на плъхове през пренаталния период на радиация, разпространяваща се от мобилни телефони, върху бъбречното развитие
Принадлежности- PMID: 25691088
- DOI: 10.3109 / 0886022X.2014.985995
Резюме
Предистория: Целта на това проучване е да изследва ефектите от излагането на 900-MHz електромагнитно поле (EMF), произведено от мобилни телефони, върху бъбречното развитие на пренатални плъхове. Определени са хистопатологични промени и апоптоза в бъбреците, заедно с нивата на урея, креатинин и електролит в серума.
Методи: Проучени са общо 14 плъха на Sprague-Dawley. Бременните плъхове бяха разделени на две равни групи: контролна група и група, изложена на EMF. Изследваната група е била изложена на 900-MHz EMF през първите 20 дни от бременността, докато контролната група е била неекспонирана на EMF. Секции, получени от парафинови блокове, бяха оцветени за каспаза-3 чрез имунохистохимия, хематоксилин-еозин и трихром на Masson.
Резултати: В бъбречните тъкани на плъхове от изложената група са наблюдавани леки задръствания и тубулни дефекти и дилатация на капсулата на Bowman. Апоптозата се оценява с помощта на анти-каспаза-3; по-силно положително оцветяване се наблюдава в бъбречните тубуларни клетки в изследваната група от тези на контролната група. Въпреки че имаше значителна разлика между изследваните и контролните групи по отношение на нивото на K + (p <0,05), не се наблюдава значителна разлика в другите изследвани параметри (p> 0,05).
Заключение: Нашето проучване показва, че електромагнитните вълни, разпространявани от мобилни телефони, имат вредно въздействие върху бъбречното развитие на пренатални плъхове.
Ключови думи: Каспаза-3; бъбреци; мобилни телефони; плъх.
Подобни статии
- Ефектите от пренаталната експозиция на 900-MHz електромагнитно поле върху 21-дневното сърце на мъжки плъх.Electromagn Biol Med. 2015; 34 (4): 390-7. doi: 10.3109 / 15368378.2014.952742. Epub 2014 28 август.PMID: 25166431
- Ефекти от пренаталната експозиция на електромагнитно поле от 900 MHz върху хистологията на бъбреците на плъхове.Int J Radiat Biol. 2015 януари; 91 (1): 35-41. doi: 10.3109 / 09553002.2014.950436.PMID: 25084839
- Ефект на излагане и отнемане на 900-MHz електромагнитни вълни върху мозъчния, бъбречния и чернодробния оксидативен стрес и някои биохимични параметри при мъжки плъхове.Electromagn Biol Med. 2015; 34 (4): 279-84. doi: 10.3109 / 15368378.2014.906446. Epub 2014 8 април.PMID: 24712749
- Настоящо разбиране за здравните ефекти на електромагнитните полета.Педиатър Ан. 2017 1 април; 46 (4): e172-e174. doi: 10.3928 / 19382359-20170316-01.PMID: 28414399 Преглед.
- [Ефекти на електромагнитните полета, генерирани от мобилните телефони върху нервната система].Med Pr. 2004; 55 (5): 435-8.PMID: 15768897 Преглед. Полски.
Цитирано от 3 статии
- Ефект на 900-MHz електромагнитно поле върху малкия мозък: Хистопатологично изследване.Sisli Etfal Hastan Tip Bul. 2018 юни 1; 52 (2): 129-134. doi: 10.14744 / SEMB.2018.42275. eCollection 2018.PMID: 32595386 Безплатна статия от PMC.
- Ефекти на фолиевата киселина върху бъбреците на плъхове, изложени на 900 MHz електромагнитно излъчване.J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 198-205. doi: 10.1016 / j.jmau.2017.06.001. Epub 2017 17 юни.PMID: 30023255 Безплатна статия от PMC.
- Грижи преди зачеването: нов стандарт на грижи в рамките на здравните услуги за майките.Biomed Res Int. 2016; 2016: 6150976. doi: 10.1155 / 2016/6150976. Epub 2016 29 май.PMID: 27314031 Безплатна статия от PMC.
Дозиметрично изследване на експозицията на плода на еднородни магнитни полета при 50 Hz. Liorni, et al. Биоелектромагнетика 35 (8): 580-97 (2014).Биоелектромагнетика
doi: 10.1002 / bem.21878. Epub 2014 29 септември.. 2014 декември; 35 (8): 580-97.Дозиметрично изследване на излагане на плода на еднородни магнитни полета при 50 Hz
Принадлежности- PMID: 25266786
- DOI: 10.1002 / bem.21878
Резюме
В тази статия излагането на плода на еднородни магнитни полета (MF) с различни поляризации се определя количествено при 50 Hz. Бяха извършени цифрови изчисления на бременни модели с висока резолюция на 3, 7 и 9 месеца от гестационната възраст (GA), които отличават голям брой фетални тъкани. Феталните индуцирани от цялото тяло и тъканите електрически полета (E) и плътности на тока (J) бяха анализирани като функция както на поляризацията на изключително нискочестотното магнитно поле (ELF-MF), така и на GA. Освен това, индуцираната вариация на полето поради промени в позицията на плода беше анализирана с помощта на два нови бременни модела. Бюджетът на несигурността поради разделителната способност на мрежата също беше изчислен. Накрая беше проверено съответствието на експозицията на плода с насоките на ICNIRP. Изградена е матрица за експозиция на плода при 50 Hz, което би могло да се използва за допълнително изследване на възможните механизми на взаимодействие между ELF-MF и свързания с това риск за здравето. Някои специфични открития са: (1) индуцираните полета се увеличават с GA; (2) максимумите Е са открити в кожни и мастни тъкани при всеки GA; (3) феталната тъкан-специфична експозиция е модифицирана като функция на GA и поляризация; (4) промяната на положението на плода в утробата значително променя индуцирания Е в някои фетални тъкани; (5) индуцираните полета бяха в съответствие с насоките на ICNIRP и резултатите бяха доста под допустимата прагова граница. (3) феталната тъкан-специфична експозиция е модифицирана като функция на GA и поляризация; (4) промяната на положението на плода в утробата значително променя индуцирания Е в някои фетални тъкани; (5) индуцираните полета бяха в съответствие с насоките на ICNIRP и резултатите бяха доста под допустимата прагова граница. (3) феталната тъкан-специфична експозиция е модифицирана като функция на GA и поляризация; (4) промяната на положението на плода в утробата значително променя индуцирания Е в някои фетални тъкани; (5) индуцираните полета бяха в съответствие с насоките на ICNIRP и резултатите бяха доста под допустимата прагова граница.
Ключови думи: ELF магнитни полета; числена дозиметрия; излагане на бременна жена.
© 2014 Wiley Periodicals, Inc.
Коментирайте в
- Коментари за "Дозиметрично изследване на излагането на плода на еднородни магнитни полета при 50 Hz" от Liorni et al.Биоелектромагнетика. 2015 април; 36 (3): 244. doi: 10.1002 / bem.21905. Epub 2015 12 март.PMID: 25763806 Няма наличен резюме.
Подобни статии
- КОМПЮТЪРНА ОЦЕНКА НА БРЕМЕННИ ЖЕНСКИ МОДЕЛИ, ИЗЛОЖЕНИ В ЕДИННИ ЕЛЕМАГНИТНИ ПОЛЯ: СЪОТВЕТСТВИЕ С ЕВРОПЕЙСКИТЕ ТЕКУЩИ РЕГЛАМЕНТИ ЗА ЕКСПОЗИЦИЯ ЗА ОБЩИТЕ ОБЩЕСТВЕНИ И ПРОФЕСИОНАЛНИ ЕКСПОЗИЦИИ НА 50 Hz.Дозиметрия на Radiat Prot. 2016 декември; 172 (4): 382-392. doi: 10.1093 / rpd / ncv488. Epub 2015 30 ноември.PMID: 26628611
- Излагане на плодове с висока разделителна способност при напреднали модели на бременни жени на различни етапи от бременността на еднородни магнитни полета с честота 50 Hz.Annu Int Conf IEEE Eng Med Biol Soc. 2013; 2013: 4525-8. doi: 10.1109 / EMBC.2013.6610553.PMID: 24110740
- Числена дозиметрия ELF: точност на метода, променливост на модели и параметри и значение за количествените насоки.Здраве Физ. 2007 юни; 92 (6): 521-30. doi: 10.1097 / 01.HP.0000251249.00507.ca.PMID: 17495652 Преглед.
- Анализ на излагането на човешкия мозък на нискочестотни магнитни полета: числена оценка на пространствено осреднени електрически полета и граници на експозиция.Биоелектромагнетика. 2013 юли; 34 (5): 375-84. doi: 10.1002 / bem.21780. Epub 2013 12 февруари.PMID: 23404214
- Оценка на биологичните ефекти, дозиметрични модели и оценка на експозицията, свързани с ELF насоките за електрическо и магнитно поле.Appl Occup Environ Hyg. 2001 декември; 16 (12): 1118-38. doi: 10.1080 / 10473220127412.PMID: 11783873 Преглед.
Цитирано от 5 статии
- Влияние на проводимостта на тъканите върху излагането на плода на изключително нискочестотни магнитни полета при 50 Hz, използвайки стохастична дозиметрия.PLoS One. 2018 7 февруари; 13 (2): e0192131. doi: 10.1371 / journal.pone.0192131. eCollection 2018.PMID: 29415005 Безплатна статия от PMC.
- Стохастична дозиметрия за оценка на излагането на деца на равномерно магнитно поле 50 Hz с несигурна ориентация.Biomed Res Int. 2017; 2017: 4672124. doi: 10.1155 / 2017/4672124. Epub 2017 31 октомври.PMID: 29226139 Безплатна статия от PMC.
- Грижи преди зачеването: нов стандарт на грижи в рамките на здравните услуги за майките.Biomed Res Int. 2016; 2016: 6150976. doi: 10.1155 / 2016/6150976. Epub 2016 29 май.PMID: 27314031 Безплатна статия от PMC. Преглед.
- Изследване на влиянието на ориентацията на 50-Hz магнитно поле върху експозицията на плода, използвайки полиномно разлагане на хаоса.Int J Environment Res Public Health. 2015 г. 27 май; 12 (6): 5934-53. doi: 10.3390 / ijerph120605934.PMID: 26024363 Безплатна статия от PMC.
- Оценка на излагането на плода на хомогенния хармоничен спектър на магнитното поле, генериран от електропреносните и разпределителните мрежи.Int J Environment Res Public Health. 2015 1 април; 12 (4): 3667-90. doi: 10.3390 / ijerph120403667.PMID: 25837346 Безплатна статия от PMC.
Влияние на етапа на бременността и позицията на плода върху цялото тяло и локалната експозиция на плода на RF-EMF. Varsier, et al. Физика в медицината и биологията 59 (17): 4913-26 (2014).
Влияние на етапа на бременността и положението на плода върху цялото тяло и локалната експозиция на плода на RF-EMF
Принадлежности- PMID: 25098501
- DOI: 10.1088 / 0031-9155 / 59/17/4913
Резюме
Тази статия анализира влиянието на етапа на бременността и позицията на плода върху излагането на цялото тяло и мозъка на плода на радиочестотни електромагнитни полета. Нашият анализ се извършва с помощта на полухомогенни модели на бременни жени между 8 и 32 седмици на аменорея. Като анализираме влиянието на етапа на бременността върху околната среда на цялото тяло и локалната експозиция на плода във вертикално положение, с главата надолу или с главата нагоре, в честотната лента 2100 MHz, стигнахме до извода, че експозицията на цялото тяло и средната мозъчна експозиция на плодът намалява през първия триместър на бременността, докато те напредват по време на бременността поради бързото наддаване на тегло на плода през тези първи етапи. От началото на втория триместър, експозициите на цялото тяло и средните мозъци са доста стабилни, тъй като увеличаването на теглото е почти пропорционално на увеличената абсорбирана мощност. Установено е, че поведението на цялото тяло на плода и локални експозиции по време на бременност за плода във вертикално положение с главата нагоре са на подобно ниво, в сравнение с положението с главата надолу те са малко по-високи, особено в мозък.
Подобни статии
- Излагане на RF-EMF на плода и майката по време на ядрено-магнитен резонанс.Phys Med Biol. 2008 декември 21; 53 (24): 7187-95. doi: 10.1088 / 0031-9155 / 53/24/012. Epub 2008 26 ноември.PMID: 19033645
- Не са открити неблагоприятни ефекти при едновременно излагане на цялото тяло на многочестотни радиочестотни електромагнитни полета за плъхове през вътрематочния и преди и след отбиването период.J Radiat Res. 2017 януари; 58 (1): 48-58. doi: 10.1093 / jrr / rrw085. Epub 2016 21 септември.PMID: 27694283 Безплатна статия от PMC.
- Количествено определяне на радиочестотната експозиция на плода, използвайки анатомични CAD-модели в три различни гестационни етапа.Здраве Физ. 2014 ноември; 107 (5): 369-81. doi: 10.1097 / HP.0000000000000129.PMID: 25271926
- Инструменти за оценка и измерване на лични и екологични експозиции на радиочестотно-електромагнитно поле.Australas Phys Eng Sci Med. 2016 март; 39 (1): 29-42. doi: 10.1007 / s13246-015-0412-z. Epub 2015 18 дек.PMID: 26684750 Преглед.
- Излагане на ниско ниво на радиочестотни електромагнитни полета: ефекти върху здравето и нужди от изследвания.Биоелектромагнетика. 1998; 19 (1): 1-19.PMID: 9453702 Преглед.
Цитирано от 6 статии
- Асоциации на използването на мобилен телефон на майката по време на бременност с продължителност на бременността и растеж на плода в 4 кохорти при раждане.Am J Epidemiol. 2019 юли 1; 188 (7): 1270-1280. doi: 10.1093 / aje / kwz092.PMID: 30995291 Безплатна статия от PMC.
- Промяна на адаптивното поведение на потомството след излагане на майчин мобилен телефон.Environment Sci Pollut Res Int. 2018 април; 25 (11): 10894-10903. doi: 10.1007 / s11356-017-1178-5. Epub 2018 3 февруари.PMID: 29397508
- Използване на мобилен телефон на майката в началото на бременността и езика, комуникацията и двигателните умения на детето на 3 и 5 години: кохортното проучване на норвежката майка и дете (MoBa).BMC обществено здраве. 2017 септември 5; 17 (1): 685. doi: 10.1186 / s12889-017-4672-2.PMID: 28870201 Безплатна статия от PMC.
- Използване на мобилен телефон на майката по време на бременност и детски поведенчески проблеми в пет кохорти при раждане.Environment Int. 2017 юли; 104: 122-131. doi: 10.1016 / j.envint.2017.03.024. Epub 2017 7 април.PMID: 28392066 Безплатна статия от PMC.
- Грижи преди зачеването: нов стандарт на грижи в рамките на здравните услуги за майките.Biomed Res Int. 2016; 2016: 6150976. doi: 10.1155 / 2016/6150976. Epub 2016 29 май.PMID: 27314031 Безплатна статия от PMC. Преглед.
Социални аномалии, свързани с аутизма, при мишки, изложени перинатално на изключително нискочестотни електромагнитни полета. Alsaeed и др. Международен вестник за неврология в развитието 37: 58-6 (2014).
Съответни на аутизма социални аномалии при мишки, изложени перинатално на изключително нискочестотни електромагнитни полета
Принадлежности- PMID: 24970316
- DOI: 10.1016 / j.ijdevneu.2014.06.010
Резюме
Честотата на разстройствата от аутистичния спектър (ASD) нараства, но причините за ASD остават до голяма степен неидентифицирани. Колективните данни предполагат повишеното излагане на човека на електромагнитни полета (ЕМП) в нарастващата честота на ASD. Има установени биологични ефекти на EMF с изключително ниска честота (ELF), но връзката с ASD не е достатъчно проучена. В това проучване изследвахме ефектите от перинаталната експозиция на ELF EMF върху някои свързани с ASD поведенчески параметри при мишки. ЕМП се доставя чрез двойка намотки Helmholtz. Бяха използвани мъжки мишки BALB / C и разделени на експонирани и контролни групи (n = 8 и n = 9, съответно). Тестовете бяха използвани за оценка на общителността, предпочитанието към социална новост, движение, тревожност, експлораторно поведение, двигателна координация и обоняние. Изследваните мишки са били мъже и са били изложени на ЕМП през последната гестационна седмица и в продължение на 7 дни след раждането. Изложените мишки демонстрират липса на нормална общителност и предпочитание към социална новост, като същевременно поддържат нормално тревожно поведение, движение, двигателна координация и обоняние. Изложените мишки също демонстрират намалена изследователска активност. Заключихме, че тези резултати подкрепят хипотезата за причинно-следствена връзка между излагане на ELF-EMF и ASD; препоръчват се обаче повторения на изследването с допълнителни тестове. Изложените мишки също демонстрират намалена изследователска активност. Заключихме, че тези резултати подкрепят хипотезата за причинно-следствена връзка между излагане на ELF-EMF и ASD; препоръчват се обаче повторения на изследването с допълнителни тестове. Изложените мишки също демонстрират намалена изследователска активност. Заключихме, че тези резултати подкрепят хипотезата за причинно-следствена връзка между експозицията на ELF-EMF и ASD; препоръчват се обаче повторения на изследването с допълнителни тестове.
Ключови думи: безпокойство; Разстройство от аутистичния спектър; Електромагнитно поле; Моторна координация; Перинатална експозиция; Социална новост.
Авторско право © 2014 ISDN. Публикувано от Elsevier Ltd. Всички права запазени.
Подобни статии
- Физическата активност като опция за намаляване на неблагоприятния ефект от излагането на ЕМП по време на бременност.Int J Dev Neurosci. 2018 декември; 71: 10-17. doi: 10.1016 / j.ijdevneu.2018.07.009. Epub 2018 1 август.PMID: 30076989
- Ефекти от излагането на изключително нискочестотни електромагнитни полета върху пространственото и пасивно избягване на ученето и паметта, подобно на тревожност поведение и оксидативен стрес при мъжки плъхове.Behav Brain Res. 2019 февруари 1; 359: 630-638. doi: 10.1016 / j.bbr.2018.10.002. Epub 2018 2 октомври.PMID: 30290199
- Краткосрочни ефекти от експозицията на електромагнитни полета с изключително ниска честота върху болестта на Алцхаймер при плъхове.Int J Radiat Biol. 2015 януари; 91 (1): 28-34. doi: 10.3109 / 09553002.2014.954058. Epub 2014 14 ноември.PMID: 25118893
- Насоки EUROPAEM EMF 2016 за профилактика, диагностика и лечение на здравни проблеми и заболявания, свързани с ЕМП.Rev Environment Health. 2016 г. 1 септември; 31 (3): 363-97. doi: 10.1515 / reveh-2016-0011.PMID: 27454111 Преглед.
- Изключително нискочестотни електромагнитни полета като ефектори на клетъчни реакции in vitro: възможно активиране на имунните клетки.J Cell Biochem. 2004 1 септември; 93 (1): 83-92. doi: 10.1002 / jcb.20198.PMID: 15352165 Преглед.
Цитирано от 4 статии
- Зъбните пломби от амалгама и използването на технологични устройства като фактор на околната среда: Актуализиране на кумулативната хипотеза за аутизъм, базирана на експозиция на живак.Eur J Dent. 2017 октомври-декември; 11 (4): 569-570. doi: 10.4103 / ejd.ejd_222_17.PMID: 29279688 Безплатна статия от PMC. Няма наличен резюме.
- Грижи преди зачеването: нов стандарт на грижи в рамките на здравните услуги за майките.Biomed Res Int. 2016; 2016: 6150976. doi: 10.1155 / 2016/6150976. Epub 2016 29 май.PMID: 27314031 Безплатна статия от PMC. Преглед.
- Ефекти от изключително нискочестотни електромагнитни полета (100 μT) върху поведението при плъхове.Невротоксикология. 2016 януари; 52: 104-13. doi: 10.1016 / j.neuro.2015.11.010. Epub 2015 22 ноември.PMID: 26593281 Безплатна статия от PMC.
- Обединяване на данни от генетични и епигенетични подходи за по-добро разбиране на разстройството на аутистичния спектър.Епигеномика. 2016 януари; 8 (1): 85-104. doi: 10.2217 / epi.15.92. Epub 2015 9 ноември.PMID: 26551091 Безплатна статия от PMC.
Пирамидална клетъчна загуба в Cornu Ammonis на 32-дневни женски плъхове след излагане на 900 мегагерцово електромагнитно поле през пренаталните дни 13–21. Bas, et al. NeuroQuantology том 11, брой 4: 591-599 (2013).
NeuroQuantology

Издател : AnKa Publisher
Редакция: Поглед в бъдещето и отворена покана за научна общност
Това, което наричаме наука, е систематизирането на информацията, получена от природата. Природата има свои собствени закони от самото начало. Някои от тези закони са лесни за изразяване, докато други разширяват нашето разбиране и дори нашето чувство за логика. Усилията ни да разберем природата и нейното функциониране, което е нашето производство на научно познание, никога няма да свършат. Може никога да не разберем истински действията на природата или да се доближим до истинската истина. Следователно, смешно е да се държим така, сякаш познаваме всички действия на природата и да кажем, че това не е научно, а е в противоречие с (известните) закони на науката ??. Най-яркият пример за това е, когато виждаме работата на квантовата физика в биологичните структури. Когато природата работи, тя не познава законите на нашата наука и дори не ги забелязва. Природата дори понякога ни намига с аномалии ??. Учим се от природата, но не можем да наложим на природата законите, които сме научили от нея.
Ефектите от 900 мегагерцови електромагнитни полета, приложени в пренаталния период върху морфологията на гръбначния мозък и двигателното поведение при женски плъхове. Odaci и др. NeuroQuantology том 11, брой 4: 573-581 (2013).
Излагане на радиочестотна радиация на плода от клетъчни телефони с номинална честота 800-1900 MHz влияе върху невроразвитието и поведението на мишките. Aldad, Gan, et al. Научни доклади 2 (312) (2013).
Излагането на радиочестотна радиационна радиация от клетъчни телефони с номинална честота от 800 MHz до Mhz влияе върху невроразвитието и поведението на мишките
Научни доклади 2 , Номер на : 312 ( 2012 )
Резюме
Невроповеденческите разстройства са все по-разпространени при децата, но тяхната етиология не е добре разбрана. Постулира се връзката между пренаталната употреба на клетъчен телефон и хиперактивността при деца, но преките ефекти на излагането на радиочестотна радиация върху невроразвитието остават неизвестни. Тук използвахме модел на мишка, за да покажем, че вътреутробното радиочестотно излагане от клетъчни телефони влияе върху поведението на възрастните. Мишки, изложени вътреутробноса били хиперактивни и са имали увредена памет, както е определено с помощта на разпознаване на обекти, светлина / тъмна кутия и понижаващи тестове. Записите със скоби на целите клетки на миниатюрни възбуждащи постсинаптични токове (mEPSC) разкриват, че тези поведенчески промени се дължат на променено програмиране на развитието на невроните. Изложените мишки са имали дозово-чувствително нарушено глутаматергично синаптично предаване върху пирамидални неврони от слой V на префронталната кора. Представяме първите експериментални доказателства за невропатология, дължаща се на вътреутробно клетъчно излъчване. Необходими са допълнителни експерименти при хора или примати, които не са хора, за да се определи рискът от експозиция по време на бременност.
Въведение
Към днешна дата 3–7% от децата в училищна възраст страдат от хиперактивно разстройство с дефицит на вниманието (ADHD) 1 . Децата с диагноза ADHD са изложени на по-голям риск от ниски академични постижения, лоши училищни постижения и престъпно поведение, несъвместимо с тяхното ниво на развитие 2 , 3 . Диагнозата на ADHD се е увеличила средно с 3% годишно от 1997 г., което прави състоянието нарастваща загриженост за общественото здраве 1 . Поведенческите проблеми при ADHD са свързани с невропатология, локализирана предимно в префронталната кора. Децата с ADHD имат намаляване на обема на префронталната кора, намаляване на сивото и бялото вещество и асиметрия 4 , 5. Тези деца също имат дефицит в работната памет, свързан с невнимание и контролиран от активността на невроните в префронталната кора 6 . Едно скорошно проучване показва, че лошото внимание и капацитет ниска работна памет може да се дължи на невъзможността да се замени принудително улавяне на вниманието от ненужна информация 7 . Това също се контролира от префронталната кора, тъй като преместването на вниманието на човека доброволно се задвижва от сигнали „отгоре надолу“ в префронталната кора, докато неволното улавяне на вниманието зависи от сигналите „отдолу-нагоре“ както от подкорковите структури, така и от визуалната кора 7 .
Етиологията на ADHD остава неизвестна и нарастващите доказателства предполагат, че тя не се дължи единствено на генетични фактори 8 . Рисковите фактори включват семейна психиатрична история, социално-икономически статус, пол и пушене по време на бременност 9 , 10 . Едно скорошно проучване установи, епидемиологична връзка между пренатална клетъчен телефон експозиция и следващите поведенчески проблеми в откритата поколението 11 . Тази асоциация е важна предвид нарастващия брой потребители на клетъчни телефони по света, достигайки приблизително четири милиарда към декември 2008 г. 12 . Липсват обаче доказателства за пряка причинно-следствена връзка.
Специфичната скорост на абсорбция (SAR) е мярка за излагане на тъканна радиация. Европейският съюз е определил граница на SAR от 2,0 W / kg, а в САЩ тази граница е 1,6 W / kg 13 . На интраутеринно последиците от излагане на радиация в този срок SAR развитието на нервната система остават неизвестни. За да определим дали пренаталното излагане на радиочестотна радиация води до нарушена памет или поведение след раждането, извършихме поведенчески и електрофизиологични проучвания при мишки, изложени вътреутробно на 800–1900 Mhz радиочестотно лъчение от клетъчни телефони.
Резултати
За да определим дали излагането на радиация на вътреутробния клетъчен телефон влияе на поведението, избрахме да проведем набор от тестове, които идентифицират увреждания в паметта, хиперактивност, тревожност и страх, които често са свързани с ADHD. Тридесет и три женски мишки бяха изложени по време на бременността (дни 1–17) на излъчване от заглушени и заглушени 800–1900 Mhz клетъчни телефони с SAR от 1,6 W / kg. Телефоните бяха разположени над всяка клетка над зоната на бутилката за хранене на разстояние 4,5–22,3 cm от всяка мишка, в зависимост от местоположението на животното в клетката и бяха поставени на непрекъснат активен разговор по време на изпитанието. Контролна група от четиридесет и две женски мишки беше държана едновременно при същите условия, но използвайки деактивиран телефон. Раждането не се различава между групите и се случва на 19 дни ± 1 ден. За да се оцени паметта при експонираните и неекспонираните мишки, 161 потомство получиха стандартен тест за памет за разпознаване на обекти в три различни кохорти на 8, 12 и 16 седмична възраст (82 експериментални и 79 контролни мишки). На мишките беше позволено да изследват два еднакви обекта за 15 минути на ден в продължение на два дни и на третия ден един обект беше заменен с нов обект. На 3 ден мишките бяха заснети в продължение на 5 минути, за да изследват романа и познатите обекти. Трима наблюдатели, заслепени за лечението, разгледаха кадрите и записаха времето за проучване на новите и познати обекти. Индексът на предпочитанията се определя като времето, прекарано в проучване на новия обект, разделено на времето, прекарано в проучване както на новия, така и на стария обект, умножено по сто. Намаляването на индекса на предпочитанията показва намалена памет.Фигура 1 ]. Средният индекс на предпочитание в изложената група е 56,8, 69,4 и 63,5 в сравнение с 66,5, 71,7 и 71,2 в контролната група, съответно на 8, 12 и 16 седмици. Експерименталната група имаше кумулативен среден индекс на предпочитание от 63,0%, а контролната група 69,9% (p = 0,003, n = 161, t тест). В сравнение с контролната група, изложените мишки са имали значително по-нисък среден индекс на предпочитание, което предполага увреждане на паметта [ Фигура 1]. За да гарантираме, че нашите открития всъщност се дължат на дефицит на паметта, а не на разсеяност или хиперактивност, изчислихме процента прекарано време на празен ход - без да изследваме нито един от обектите. Средното време на празен ход в изложената група е 90,06, 90,53 и 96,48 в сравнение с 92,12, 91,89 и 97,07 в контролната група, съответно на 8, 12 и 16 седмици. Контролната група имаше кумулативно средно време на празен ход от 90,8%, докато експерименталната група имаше кумулативно средно време на празен ход от 90,4% и разликата между двете групи не беше статистически значима (p = .58).
Фигура 1 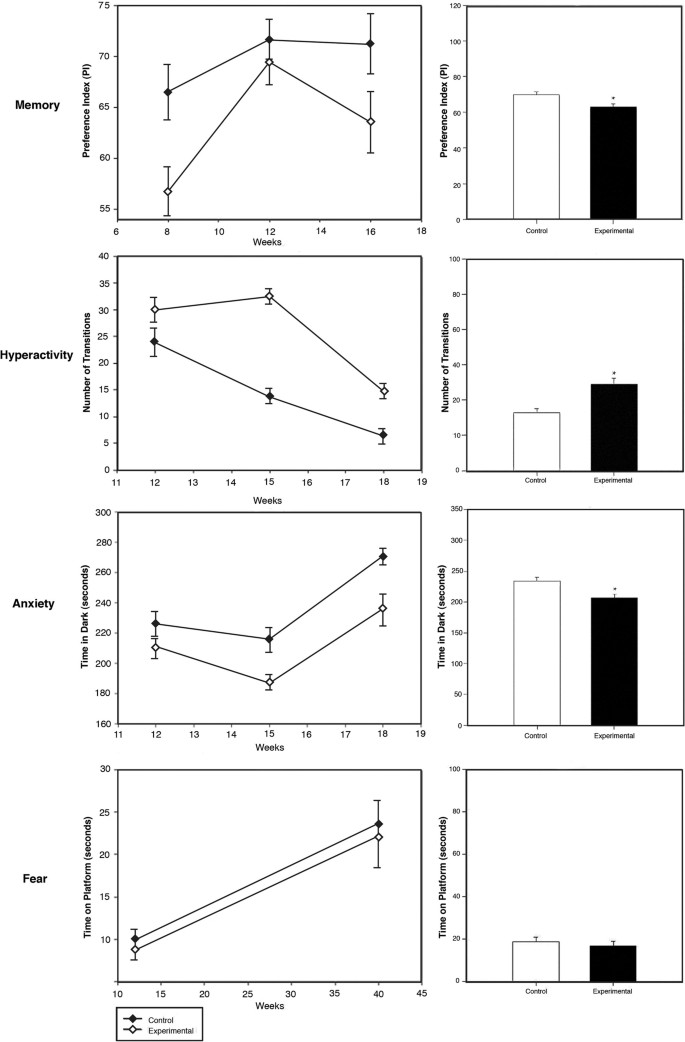
Тестове за поведение при изложени и контролни мишки.
Лявата колона показва данните, определени при мишки на няколко възрасти след излагане. В дясната колона е показана кумулативната средна стойност. За тестване на паметта беше използван стандартният тест за памет за разпознаване на обекти и показан Индекс за предпочитание (процент от общото време за изследване, прекарано в проучване на новия обект) на 8, 12 и 16 седмична възраст. Кумулативният среден индекс на предпочитание на експерименталната група е 63,0%, а на контролната група 69,9% (* р = 0,003, n = 161). За да тестваме хиперактивност, използвахме теста Light / Dark box и показахме преходи на възраст 12, 15 и 18 седмици. Кумулативният среден брой преходи в експерименталната група е 24,4, а контролната група 16,4 (* p = <0,001, n = 141). За да тестваме безпокойството, измерихме времето, прекарано на тъмно на 12, 15 и 18 седмична възраст. Кумулативното средно време, прекарано на тъмно в експерименталната група, е 207 секунди, а в контролата е 234 секунди (* р <0,001, n = 141). За измерване на страха използвахме теста Step down и показахме времето, прекарано на платформата на 12 седмици и зряла възраст. Кумулативното средно време, прекарано на платформата в експерименталната група, е 16,7 секунди, а в контролата - 18,5 секунди (p = 0,59, n = 98).
За да изследваме страховитото поведение, извършихме тест за светлина / тъмна кутия, измерващ хиперактивност / тревожност и анализ за понижаване, оценяващ страха от изследване на околната среда. Тревогата на мерки за изпитване кутия светлина / тъмнина чрез използване на природни антипатия гризач да ярка светлина 14 . Кутията съдържаше две отделения: едно бяло отделение, което беше осветено и едно черно отделение, което остана тъмно. Броят на преходите между двете отделения е използван за определяне на движението и от своя страна хиперактивност 15 . Тревожното поведение се измерва чрез записване на времето, прекарано във всяко отделение 15. На общо 141 потомство е направен тест светлина / тъмна кутия в три различни кохорти на възраст 12, 15 и 18 седмици (71 експериментални и 70 контролни мишки). Всяка мишка беше поставена в светло / тъмно поле за 5 минути и заснета. Трима наблюдатели, заслепени за режима на лечение, разгледаха кадрите и записаха времето, прекарано в тъмното отделение, заедно с броя на преходите между всяко отделение. Средният брой преходи в експерименталната група на 12, 15 и 18 седмици е бил по-малък, отколкото в съответните контроли и резултатите са били значими във всеки момент от времето [ Фигура 1]. Средният брой преходи при изложени мишки е 29,9, 32,5 и 14,8 в сравнение с 23,9, 13,8 и 6,5 в контролната група, съответно на 12, 15 и 18 седмици. Експерименталната група показа кумулативна средна стойност от 24,4 прехода, а контролната група показа средна стойност от 16,4 прехода (р <0,001). В сравнение с контролната група, по-големият брой преходи между двете отделения в експерименталната група предполага хиперактивно поведение [ Фигура 1 ].
За да установим дали тревожността може да е фактор, допринасящ за поведенческия фенотип, докладван в експеримента светлина / тъмна кутия, първо сравнихме продължителността на времето в тъмното спрямо времето, прекарано в светлината. Увеличеното време на тъмно показва тревожно поведение 15 . На 12, 15 и 18 седмици експерименталната група прекарва по-малко време на тъмно и резултатите са значими във всеки момент от времето [ Фигура 1 ]. Продължителността на времето в тъмнина на изложената група е 210,8, 187,0 и 235,8 секунди в сравнение с 225,6, 215,5 и 270,6 секунди в контролната група, съответно на 12, 15 и 18 седмици. Мишките, изложени вътреутробнопрекара кумулативно средно 207 секунди на тъмно, докато контролните мишки прекараха средно 234 секунди на тъмно, което показва намалена тревожност при мишки, изложени на клетъчен телефон (p <0,001) [ Фигура 1 ].
Анализът Step Down е извършен на 98 мишки на 12 седмици и в зряла възраст, за да се определи страхът от изследване на околната среда (51 контролни и 47 експериментални мишки). Тестът се извършва чрез записване на времето, прекарано на стандартна платформа. По-големият период от време на платформата показва повишена страхливост. Изложените мишки не показаха значителна разлика във времето, прекарано на платформата в сравнение с контролите [ Фигура 1 ]. Контролните мишки са прекарали средно 18,5 секунди, докато експерименталната група е прекарала средно 16,7 секунди (p = 0,59) [ Фигура 1 ].
Като цяло мишките, изложени вътреутробно на радиация, са били хиперактивни, са имали намалена памет и намалена тревожност.
За да разберем механизмите, лежащи в основата на промените в паметта и хиперактивността при животни, изложени на радиация вътреутробно , ние изследвахме дали са настъпили промени в невронните вериги в мозъчните области, отговорни за това компрометирано поведение. По-конкретно, попитахме дали промените в синаптичното предаване в невроните на ЦНС са отговорни за нарушената памет и хиперактивността при изложени на радиация животни. Префронталната кора (PFC) е отговорна за изпълнителните функции чрез скрининг на разсейването и поддържане на вниманието в целенасочено поведение. Нарушението на PFC води до нерегулирано поведение / емоция като ADHD 16. Пирамидалните неврони, основният клетъчен тип в тази структура, регулират вниманието и поведението чрез сложна и взаимосвързана мрежа. Записите със скоби на цели клетъчни пластири на миниатюрни възбуждащи постсинаптични токове (mEPSC) се извършват в пирамидални неврони на PFC в контролни и изложени на клетъчни телефони мишки. mEPSCs се генерират чрез произволно освобождаване на глутамат от везикули от пресинаптичните неврони при липса на стимулация. Измерването на mEPSC се използва за анализ на ефикасността на синаптичното предаване. Смята се, че промените в mEPSC честотата са резултат от модификация на пресинаптичния компонент на синаптичното предаване, докато промените в амплитудата показват промени в постсинаптичния компонент 17 , 18. Коронални префронтални резени на кора (300 μm) бяха приготвени от мишки на възраст 3-4 седмици. mEPSC са регистрирани в пирамидални неврони от слой V в префронталната кора на мишки, изложени на вътреутробно облъчване в продължение на 9, 15 и 24 часа / ден през цялата бременност; откриването и анализът на mEPSC честота и амплитуда бяха извършени, както описахме по-рано 18 . При животни, изложени на вътреутробна радиация в продължение на 24 часа / ден, се наблюдава намаляване на честотата на mEPSCs (контрол: 1,00 ± 0,12 Hz, n = 40; 24 часа / ден: 0,72 ± 0,06 Hz, n = 43, p <0,05, t тест, Фигура 2А и В ). Кривите на кумулативната вероятност за амплитудата на mEPSC събития, записани от вътреутробноизложени на клетъчен телефон мишки (24 часа / ден) се изместват значително наляво спрямо тези, записани от контролите (P <0,01, тест на Колмогоров-Смирнов; контрол: 2765 събития, експозиция на мобилен телефон: 2224 събития), което показва, че на mEPSCs е намалена [ Фигура 2С ]. В подгрупа от експерименти ние изследвахме дали намаляването на честотата на mEPSC зависи от дозите на експозиция при мишки, изложени преди раждането 0, 9, 15 и 24 часа на ден [ Фигура 3 ]. Тенденцията на дозозависимото намаляване на честотата на mEPSC (0 час / ден: 1,37 ± 0,41, n = 9; 9 часа / ден: 1,27 ± 0,21 Hz, n = 9; 15 часа / ден: 1,04 ± 0,20 Hz , n = 10; 24 часа / ден: 0,72 ± 0,13, n = 11) е статистически значима (линейна корелация: Корелационен коефициент = -0,97, Некорректиран r 2 = 0,94, Р <0,05).
Фигура 2 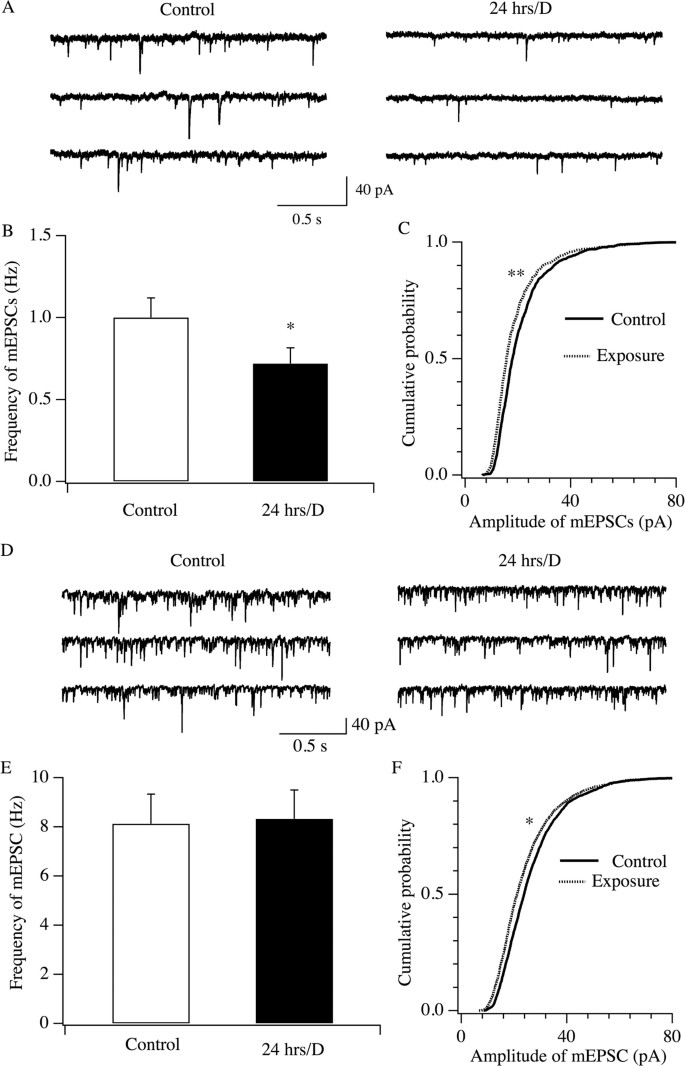
Синаптичната ефикасност на глутаматергичните синапси е намалена в мозъчните неврони на мишки след пренатално излагане на лъчение на клетъчен телефон.
A – C, mEPSC са записани в пирамидални неврони от слой V на префронталната кора. Представителните следи от mEPSC от контролни групи и групи за експозиция на клетъчни телефони са показани в А. mEPSC честота и кумулативна вероятност за амплитуда на mEPSC от двете групи са показани в B (*, P <0,05, t тест) и C (**, P <0,01 , Тест на Колмогоров-Смирнов; контроли, 2225 събития; изложени, 2766 събития). D – F, показани са представителни следи, честота и амплитуда на mEPSCs, записани в неврони във VMH. *, Р <0,05, тест на Колмогоров-Смирнов; Контрол: 2161 събития, Група мобилни телефони: 2261 събития.
Фигура 3 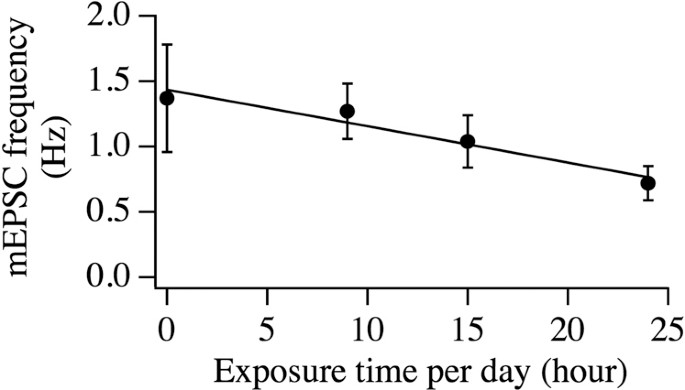
Дозозависимо затихване на честотата на mEPSCs в слой V пирамидални неврони при мишки.
Показана е честотата на mEPSCs, записани при мишки, пренатално изложени на облъчване от мобилен телефон при доза от 0, 9, 15 и 24 часа на ден. Лентите за грешки са SEM. Дозореагиращата връзка се определя с помощта на регресионен анализ (Коефициент на корелация, -0,97; r 2 , 0,94; P <0,05).
Като цяло тези резултати показват, че синаптичната ефикасност на глутаматергичното предаване намалява както на пре-, така и на постсинаптичните места в пирамидални неврони от слой V. По този начин ние демонстрираме увреждане на глутаматергичното предаване (освобождаване от нервни терминали и отговор на глутаматния рецептор) върху пирамидални неврони в PFC след вътреутробно излагане на радиация от клетъчни телефони.
В паралелен експеримент изследвахме дали вътреутробното излагане на радиация води до промени в синаптичното предаване в друга мозъчна област. mEPSC са регистрирани в неврони във вентралния медиален хипоталамус (VMH), мозъчна област, замесена в регулирането на енергийната хомеостаза 19 , 20 . Нашите резултати показват, че при мишки, изложени на радиация в продължение на 24 часа / ден, честотата на mEPSC (контрол: 8,13 ± 1,20 Hz, n = 14; излъчване на мобилен телефон: 8,32 ± 1,17 Hz, n = 14) не се различава значително от тази при контролни мишки (P> 0,05, t тест, Фигура 2D и E). Въпреки това кумулативната вероятност за амплитуда на mEPSC, записана при изложени на радиация мишки, значително се измества вляво (P <0,05, тест на Колмогоров-Смирнов; контрол: 2161 събития, група мобилни телефони: 2261 събития; Фигура 2F ), което предполага, че амплитудата на mEPSC е по-малък в групата, изложена на мобилен телефон, отколкото в контролите. Този резултат предполага, че на постсинаптичното място се получава увреждане на глутаматергичното предаване. В обобщение, нашите резултати показват, че ефектите от пренаталното излагане на лъчението на клетъчния телефон не са били ограничени до кората.
Стресът на майката може да промени развитието на плода чрез увеличаване на излагането на потомството на кортикостерон, причинявайки когнитивни дефицити, хиперактивност и промени в оста на хипоталамо-хипофизарно-надбъбречната жлеза 21 . За да се изключи възможността нарушената памет и поведение при изложени мишки да са причинени от стрес в резултат на експериментална манипулация, ние измерихме нивата на серумния кортикостерон на дванадесетия ден от бременността, използвайки ELISA анализ. Средното ниво на кортикостерон при изложени женски мишки (69,91 ng / ml, n = 6) не се различава значително от това при контролните жени (69,94 ng / ml, n = 6) [ Фигура 4 ], премахвайки стреса като източник на наблюдаваните поведенчески и електрофизиологични разлики.
Фигура 4 
Нивата на кортикостерон по време на бременност не се променят от експозицията.
Средното ниво на кортикостерон при бременните контролни жени е 69,94 ng / ml, а при изложените женски мишки е 69,91 ng / ml.
Дискусия
Тук ние демонстрираме, че излагането на плода на радиочестотна радиация от 800–1900 Mhz от клетъчни телефони води до поведенчески и неврофизиологични промени, които продължават и в зряла възраст. Мишките, изложени по време на бременност, са имали нарушена памет, били са хиперактивни и са имали намалена тревожност, което показва, че вътреутробната експозиция на радиочестота е потенциална причина за невроповеденчески разстройства. По-нататък демонстрирахме увреждане на глутаматергичното синаптично предаване върху пирамидални клетки в префронталната кора, свързано с тези поведенчески промени, предполагайки механизъм, чрез който излагането на вътреклетъчна клетъчна телефонна радиация може да доведе до увеличеното разпространение на невроповеденчески разстройства.
Това е първото проучване, което специално идентифицира ефектите от радиочестотната експозиция върху мишкия плод. По време на критични прозорци в неврогенезата мозъкът е податлив на многобройни обиди от околната среда; често срещаните медицински релевантни експозиции включват йонизиращо лъчение, алкохол, тютюн, наркотици и стрес. Ефектите на тези агенти зависят от дозата и времето на експозиция. Дори малките експозиции по време на периоди на неврогенеза имат по-дълбок ефект от експозицията като възрастен. Алкохолът засяга мозъчната неврогенеза, моделирането на мозъчното развитие и последващото поведение. Пушенето на майката също влияе върху развитието на плода; феталната експозиция на тютюн води до по-висока честота на поведенчески и когнитивни увреждания, включително ADHD. По същия начин пренаталното излагане на кокаин може да доведе до поведенчески разстройства. Дори пренаталният майчин стрес може да намали интелигентността и езиковите способности на потомството. Както се демонстрира от тези примери, излагането на околната среда, възникващо във феталния живот, може да доведе до постоянни неврологични дефицити. Излагането на тези обиди като възрастен не носи същите последствия. Ето защо не е изненадващо, че проучванията, излагащи възрастни животни на радиочестотна радиация, не успяха да открият подобни значими дефекти в поведението. Излагането на клетъчни телефони по време на бременност може да има съпоставим ефект върху плода и подобни последици за обществото, както и излагането на други често срещани токсични фактори за невроразвитие. Въпреки че тези данни показват ясна връзка между експозицията на плода на EMR и невроразвитието, важно е да се признае, че екстраполацията на този животински модел върху хората е ограничена;
Молекулярните и клетъчните ефекти на радиочестотната експозиция все още не са напълно характеризирани. Ин витро са идентифицирани множество цели . Доказано е, че излагането на електромагнитна честота влияе върху клетъчното делене и пролиферация, както чрез индуциране на апоптоза, така и чрез промяна на клетъчния цикъл 22 . Електромагнитното лъчение може да насърчи образуването на реактивни кислородни форми (ROS), причиняващи увреждане на клетките 23 . Едно проучване, специално анализиращо ефектите на радиочестотната радиация върху глиомните клетки, показва променен оксидативен стрес, потенциален медиатор на промените, причинени от електромагнитното лъчение 24 . Установено е също, че електромагнитното честотно излъчване активира ERK и p38 MAPK сигнализиране 25. Въпреки че точните молекулярни механизми, които са довели до променено глутаматергично синаптично предаване в префронталната кора, идентифицирани в това проучване, все още не са напълно известни, тук предоставяме първите доказателства, които свързват промените в невронните вериги, центрирани върху слой V пирамидални неврони в PFC с нарушена памет и когнитивно поведение при животни, изложени на радиация от използването на клетъчни телефони. Нашите резултати показват, че освобождаването на глутамат от нервните терминали на PFC невроните и отговорът на PFC невроните към глутамат са нарушени при мишки, които са били изложени преди раждането на клетъчен телефон. Тези резултати са в съответствие с предишни доклади, че компрометираното глутаматергично предаване върху PFC неврони е в основата на нарушената памет и когнитивните функции при животните 26 , 27. Нашите резултати също така показват, че ефектите от пренаталното излагане на радиация върху мозъка може да са глобални, тъй като глутаматергичното предаване върху неврони в друга област на мозъка (т.е. VMH) също е намалено. Ефектите от пренаталното излагане на радиация на клетъчния телефон може да имат по-дълбоки ефекти върху мозъчните функции, отколкото се съобщава в това проучване. Ефектът обаче не е идентичен; има вероятност да има специфични за региона тип или регионални вариации в чувствителността. Алтернативно, дълбочината на VMH може да е предпазила този регион от максимална експозиция.
Като се имат предвид неотдавнашните постижения в технологията на клетъчните телефони (т.е. смарт телефони), те вече се използват в капацитет извън този на основния телефон. За мнозина клетъчните телефони се използват като нощно будилник и личен органайзер. Използването на клетъчни телефони може да достигне 24 часа в денонощието, оставяйки потребителите все по-изложени на потенциално вредните ефекти от излагането на радиочестотни лъчи. Нашите открития показват значителни електрофизиологични и поведенчески промени при мишки, изложени вътреутробнодо радиация. Значителната тенденция между групите, лекувани за 0, 9, 15 и 24 часа / ден, показва, че ефектите са пряко пропорционални на времето за употреба и предполага, че могат да бъдат установени граници на безопасност, особено за бременни жени. Въпреки че е трудно да се преведат тези констатации на човешките рискове и уязвимост, ние идентифицираме нов потенциален принос за увеличеното разпространение при хиперактивни деца, който лесно се предотвратява. Важно е обаче да се отбележи, че хиперактивността и тревожността са тясно свързани и ме смущават.
В това проучване използвахме клетъчни телефони като източник на EMR, за да имитираме експозицията на човека. Въпреки това има няколко ограничения на това проучване, които включват липса на определена експозиция от традиционен генератор на ЕМП. Освен това не измерихме нивото на експозиция и разстоянието до източника не беше фиксирано; мишките бяха свободни да се придвижват в рамките на клетката. Измерванията на плътността на мощността по отношение на ориентация, поляризация, отражение и смущения не бяха взети предвид. За да се определят максималните ефекти и потенциалните рискове, свързани с експозицията, мишките са били изложени от зачеването до раждането, но развитието на мозъка на мишката е непълно при раждането и се различава от това на хората. Докато тук бяха открити неврологични ефекти, бъдещите проучвания трябва да се фокусират върху по-тясна гестационна възраст на експозиция, използвайте генератори на ЕМП, за да определите по-точно експозицията и да ограничите вариацията в разстоянието от източника. Необходими са окончателни проучвания при хора, преди да се екстраполират тези поведенчески констатации на хората.
В обобщение, ние демонстрираме, че излагането на радиочестотна радиация на плода е довело до невроповеденчески разстройства при мишки. Предполагаме, че тези открития ще подобрят нашето разбиране за етиологията на невроповеденческите разстройства. Нарастването на поведенческите разстройства в развитите страни може поне отчасти да се дължи на приноса от излагането на фетална клетъчна телефонна радиация. По-нататъшно тестване е необходимо при хора и примати, които не са хора, за да се определи дали рисковете са сходни и да се установят безопасни граници на експозиция по време на бременност.
Методи
Тестове за експозиция и поведение
В рамките на пет отделни експеримента бяха създадени общо 27 клетки за разплод, всяка от които съдържаше 3 CD-1 женски мишки и 1 CD-1 мъжка мишка (13 експериментални клетки и 14 контролни клетки). Всяка експериментална клетка беше снабдена със заглушен и заглушен клетъчен телефон 800–1900 Mhz със SAR от 1,6 W / kg, поставен върху зоната на бутилката за хранене на разстояние 4,5–22,3 cm от мишките. След това клетъчните телефони бяха включени в активен разговор за 24 часа на ден и 33-те експериментални женски мишки бяха изложени по време на бременността (дни 1-17). Допълнително шест жени са били изложени на активен телефон в продължение на 9 или 15 часа на ден. Всяка контролна клетка е оборудвана с деактивиран телефон и се поддържа при същите условия. За да се осигури еднакво време на експозиция, независимо от променливата продължителност на бременността (18–20 дни), в края на ден 17 всички телефони бяха премахнати. На 18 ден всички женски мишки бяха разделени и поставени в собствената им клетка, като се получиха общо 39 открити бременни жени и 42 неекспонирани бременни жени. По време на експеримента, както контролните, така и експерименталните мишки се хранят и им се дава вода ad libitum. Мишките бяха поддържани на 12-часов цикъл светлина / тъмнина (07:00 часа нататък) и всички процедури бяха одобрени от Комитета за грижа и употреба на животните в Йейлския университет.
Паметта беше оценена с помощта на стандартен тест за памет за разпознаване на обекти. Тествани са общо 161 малки (82 експериментални мишки и 79 контролни мишки) на 8, 12 и 16 седмици. Тестът се състоеше от два учебни дни (ден 1 и 2) и един ден за тест (ден 3). На 1-ви ден в изпитната стая бяха създадени четири непрозрачни изследователски камери със светимост 420–440 Lux. Преди провеждането на всеки тест, мишките бяха поставени в помещението за тестване и им беше оставен 1 час да се приспособят към светлината. След това във всяка от четирите камери бяха поставени два еднакви предмета и във всяка камера беше поставена по една мишка, за да изследва двата идентични обекта за 15 минути. Преди да се повтори експериментът, предметите и камерите се почистват старателно с детергент, за да се премахнат всякакви аромати или миризми. На Ден 3 видеокамера беше поставена над всичките 4 камери и обектите бяха пренаредени така, че всяка камера имаше по един познат обект и един нов обект. След това на мишките беше позволено да изследват двата обекта и бяха заснети за 5 минути. След приключване на експеримента, 3 наблюдатели, заслепени от режима на лечение, разгледаха първите 2 минути кадри, за да определят времето, прекарано в проучване на новия обект. Проучването на новия обект беше определено като подсмърчане на по-малко от 1 см. След това се изчислява индекс на предпочитание чрез разделяне на времето, прекарано в проучване на новия обект, на общото време за проучване, умножено по сто. Процентът време, прекарано на празен ход - непроучване на нито един от обектите, също беше изчислено, за да се гарантира, че нашите констатации всъщност се дължат на дефицити в паметта, а не на разсеяност или хиперактивност.
Тестът за светлина-тъмни кутии беше проведен с помощта на светло-тъмна кутия, изградена от черно-бял плексиглас (45 × 27 × 27 cm). Тъмното отделение (18 × 27 см) е направено от черен плексиглас с черен капак от плексиглас, а светлото отделение (27 × 27 см) е направено от бял плексиглас и остава отворено. Светлинното отделение се поддържаше със светимост 420–440 Lux. В стената между двете камери беше разположен отвор (7,5 × 7,5 см), позволяващ свободен достъп между светлите и тъмните отделения. След това над камерата беше поставена видеокамера за заснемане. Преди провеждането на всеки тест, мишките бяха поставени в помещението за тестване и им беше оставен 1 час да се приспособят към светлината. След това една мишка беше поставена в светлинната камера и беше оставена да изследва кутията в продължение на 5 минути, докато беше заснет. Преди да повторите експеримента, камерите бяха почистени старателно с детергент, за да се премахнат всякакви аромати или миризми. Трима наблюдатели, заслепени за режима на лечение, след това разгледаха кадрите и записаха общото време, прекарано на тъмно, както и общия брой преходи. След това тези данни бяха интерпретирани, както е описано в текста, за да се анализира тревожността и хиперактивността.
Анализът Step Down е извършен за определяне на страховито поведение чрез поставяне на мишка внимателно върху платформа (плоча с 96 ямки) и записване на времето на платформата. Таймерът беше спрян, след като мишката излезе от платформата с четирите лапи. Преди да се повтори експериментът, платформата беше почистена старателно с детергент, за да се премахнат всички аромати или миризми.
Измерване на кортикостерон
Гестационният стрес беше анализиран чрез събиране на серум на 12-ия ден от бременността от 6 експонирани и 6 неекспонирани бременни жени. Серумните проби бяха тествани за нива на кортикостерон, като се използва ензимен имуноанализ (Assay Designs, Ann Arbor, MI), както се препоръчва от производителя.
Електрофизиология
Мишките от контролни групи и групи, изложени на мобилни телефони, се упояват с етер и след това се обезглавяват. Мозъците бяха бързо отстранени и потопени в кислороден разтвор за рязане при 4 ° C, съдържащ (в mM): захароза 220, KCl 2,5, CaCl 2 1, MgCl 2 6, NaH 2 PO 4 1,25, NaHCO 3 26 и глюкоза 10 и коригирани до рН 7,3 с NaOH. Коронални кортикални резени (с дебелина 300 µm) се приготвят от префронталната област на мозъка и вентралния медиален хипоталамус (VMH) с помощта на вибратом. След приготвянето, резените се поддържат в камера за задържане с изкуствена цереброспинална течност (ACSF) (мехурчета с 5% CO2 и 95% O2), съдържаща (в mM): NaCl 124, KCl 3, CaCl 2 2, MgCl 22, NaH 2 PO 4 1,23, NaHCO 3 26, глюкоза 10, рН 7,4 с NaOH и се прехвърлят в записваща камера, непрекъснато перфузирана с разтвор за баня (33 ° С) при 2 ml / min след най-малко 1 час възстановяване.
Цялоклетъчна скоба за напрежение (при -60 mV) беше извършена за наблюдение на миниатюрни възбуждащи постсинаптични токове (mEPSC) в кортикални неврони от слой V с усилвател Multiclamp 700 A (Molecular devices, CA). Пластмасовите пипети (съпротивление на върха = 4–6 MΩ) са направени от боросиликатно стъкло (World Precision Instruments) с издърпващо устройство за пипети (Sutter P-97) и отзад пълни с разтвор за пипета, съдържащ (в mM): K-глюконат 135 MgCl 2 2, HEPES 10, EGTA 1.1, Mg-ATP 2, Na 2 -фосфокреатин 10 и Na 2-GTP 0,3, pH 7,3 с KOH. mEPSCs бяха записани в пирамидални неврони под напрежение (при -60 mV) в присъствието на тетродотоксин (TTX, 0.5μM,) и антагонист на GABA-A рецептор пикротоксин (50 μM). Както входното съпротивление, така и серийното съпротивление се наблюдават постоянно по време на експерименти. Последователното съпротивление (между 20 и 40 MΩ) беше частично компенсирано от усилвателя и бяха приети само записи със стабилни серии и входно съпротивление по време на експериментите. Всички данни са взети на проби при 3–10 kHz и са филтрирани на 1–3 kHz с компютър Apple Macintosh с помощта на Axograph X (AxoGraph Scientific). mEPSC събития бяха открити и анализирани с AxoGraph X и начертани със софтуера Igor Pro (WaveMetrics, Lake Oswego, OR), както е описано по-рано от Rao, et al (2007). Извършена е линейна корелация със софтуера GB-STAT (Dynamic Microsystems,
История на промените
18 февруари 2013 г.
Публикувана е корекция, която е приложена към HTML и PDF версиите на тази статия. Грешката не е отстранена в хартията.
Препратки
Американска психиатрична асоциация: Диагностично и статистическо ръководство за психични разстройства (2000) Четвърто издание, Текстова ревизия. Вашингтон, Американска психиатрична асоциация.
Barkley, R. Инхибиране на поведението, устойчиво внимание и изпълнителни функции: Изграждане на обединяваща теория на ADHD. Psychol Bull 121, 65–94 (1997).
Рапли, М. Разстройство с дефицит на вниманието и хиперактивност. N Engl J Med 352, 165–73 (2005).
Sowell, ER Кортични аномалии при деца и юноши с хиперактивно разстройство с дефицит на внимание. Lancet 362, 1699–707 (2003).
Castellanos, FX Траектории на развитие на аномалии на мозъчния обем при деца и юноши с дефицит на внимание / хиперактивност. JAMA 288, 1740–48 (2002).
Castellanos, FX & Tannock, R. Невронаука на разстройство с дефицит на вниманието / хиперактивност: търсенето на ендофенотипове. Nat Rev Neurosci 3, 617–28 (2002).
Fukuda, K. & Vogel, EK Човешки вариации при заменяне на вниманието. J Neurosci 29, 8726–33 (2009).
Сингх, И. Отвъд полемиката: наука и етика на ADHD. Nat Rev Neurosci 9, 957–64 (2008).
Brassett-Harknett, A. & Butler, N. Разстройство с дефицит на вниманието / хиперактивност: преглед на етиологията и преглед на литературата, свързана с корелатите и резултатите от жизнения цикъл за мъже и жени. Clin Psychol Rev. 27,188–210 (2007).
Biederman, J. & Faraone, SV Хиперактивно разстройство с дефицит на вниманието. Lancet 366, 237–248 (2005).
Divan, H. Kheifets, L. Obel, C. & Olsen, J. Пренатално и постнатално излагане на използването на мобилен телефон и поведенчески проблеми при деца. Епидемиология 19, 523–529 (2008).
Измерване на информационното общество: Индексът за развитие на ИКТ. Международен съюз по далекосъобщения. p108. (2009).
Указания за ограничаване на експозицията на вариращи във времето електрически, магнитни и електромагнитни полета (до 300 GHz). Международна комисия за защита от нейонизиращо лъчение. Здраве Физ. 74, 494-522 (1998).
Bourin, M. & Hascoet, M. Тест на светлината / тъмната кутия на мишката. Eur J Pharamocol. 463, 55–65 (2003).
Corbetta, S. et al. Хиперактивност и индуцирана от новостта хиперактивност при мишки без Rac3. Behav Brain Res 186, 246–255 (2008).
Arnsten, AF Стресови сигнални пътища, които увреждат структурата и функцията на префронталната кора. Nat Rev Neurosci 10, 410–22 (2009).
Ungless, MA, Whistler, JL, Malenka, RC et al. Единичната експозиция на кокаин in vivo индуцира дългосрочно потенциране в допаминовите неврони. Природата. 411, 583-587 (2001).
Rao, Y. et al. Продължителното събуждане предизвиква синаптична пластичност, зависима от опита, в невроните на миши хипокретин / орексин. J Clin Invest. 117, 4022–33 (2007).
Лопес, М. и сътр. Хипоталамусният AMPK и метаболизмът на мастните киселини медиират регулирането на енергийния баланс на щитовидната жлеза. Nat Med. 16 1001–8 (2010).
Xu, Y. et al. Сигнализирането PI3K в ядрото на вентромедиалното хипоталамус е необходимо за нормална енергийна хомеостаза. Cell Metab. 12 88–95 (2010).
Koehl, M. Lemaire, V. Le Moal, M. & Abrous, DN Възраст-зависим ефект от пренаталния стрес върху пролиферацията на хипокампални клетки при женски плъхове. Eur J Neurosci. 29 635–40 (2009).
Panagopoulos, DJ Chavdoula, ED Nezis, IP & Margaritis, LH Клетъчна смърт, предизвикана от GSM 900-MHz и DCS 1800-MHz радиация на мобилната телефония. Mutat Res. 626, 69–78 (2007).
Zmyślony, M. et al. Острото излагане на 930 MHz CW електромагнитно излъчване in vitro влияе върху нивото на реактивния кислород в лимфоцитите на плъхове, третирани с железни йони. Биоелектромагнетика. 25, 324–8 (2004).
Cao, Y. et al. 900-MHz микровълновото лъчение засилва неблагоприятните ефекти на гама-лъчите върху клетките SHG44. J Toxicol Environment Health A. 72, 727–32 (2009).
French, PW, Penny, R. Laurence, JA & McKenzie, DR Мобилни телефони, протеини от топлинен шок и рак. Диференциация 67, 93–97 (2001).
Jentsch, JD et al. Дисбиндин модулира префронтални кортикални глутаматергични вериги и функция на работната памет при мишки. Невропсихофармакология. 34, 2601–8 (2009).
Rubino, T. et al. Депресивният фенотип, предизвикан при възрастни женски плъхове от юношеска експозиция на THC, е свързан с когнитивно увреждане и променена невропластичност в префронталната кора. Neurotox Res. 15, 291–302 (2009).
Благодарности
Авторите благодарят на Arie Kaffman и Richard Hochberg за критичното четене на ръкописа и благодарят на Niil Odem, Michael Lee и Yuzhe Feng за техническата помощ и анализ на резултатите от тестовете за поведение. Поддържа се от безвъзмездни средства от EHHI и NICHD (HD052668).
Информация за автора
Принадлежности
Вноски
TSA лекува мишките, извършва поведенчески проучвания, анализира данните и пише ръкописа. GG и XBG извършват и анализират електрофизиологичните изследвания. HST проектира експеримента, анализира данните и редактира ръкописа.
Черепни и посткраниални скелетни вариации, индуцирани в миши ембриони от излъчване на мобилен телефон. Fragopoulou, Koussoulakos и др. Патофизиология 17 (3): 169-77 (2010).
Черепни и посткраниални скелетни вариации, индуцирани в миши ембриони от излъчване на мобилен телефон
Принадлежности- PMID: 19854628
- DOI: 10.1016 / j.pathophys.2009.10.002
Резюме
Това проучване се фокусира върху развитието на плода след леко ежедневно излагане на бременни мишки на близко полево електромагнитно излъчване, излъчвано от мобилен телефон. Разследването беше мотивирано от факта, че потенциално опасното електромагнитно излъчване, излъчвано от мобилните телефони, в момента представлява огромен обществен интерес. Физически сравними бременни мишки бяха изложени на радиочестотно излъчване GSM 900MHz, излъчвано от мобилен телефон. В рамките на 5 часа след раждането повечето малки са фиксирани, последвано от двойно оцветяване в тото и конвенционална парафинова хистология. Други малки са останали при майките си до никнене на зъби. Структурното развитие се оценява чрез изследване на новородени за наличие на аномалии и / или вариации в меките тъкани и скелетната анатомия. Изложени на електромагнитна радиочестота новородени, външно изследвани, показва нормален фенотип. Хистохимичните и хистологични проучвания обаче разкриват вариации в откритите плодове по отношение на контролните, касаещи осификацията на черепните кости и ребрата на гръдната клетка, както и изместването на мекелийския хрущял. Колетата, изследвани след поникването на зъбите, показват нормални фенотипове. Заключението е, че лекото излагане на радиация от мобилен телефон може да повлияе, макар и преходно, върху развитието на плода на мишката на ниво на осификация. Наблюдаваните вариации в развитието могат да бъдат обяснени чрез разглеждане на различния ембрионален произход и начин на осификация на засегнатите скелетни елементи. разкрива вариации в откритите плодове по отношение на контролните, касаещи осификацията на черепните кости и ребрата на гръдната клетка, както и изместването на мекелийския хрущял. Колетата, изследвани след никнене на зъби, показват нормални фенотипове. Заключението е, че лекото излагане на радиация от мобилен телефон може да повлияе, макар и преходно, върху развитието на плода на мишката на ниво на осификация. Наблюдаваните вариации в развитието могат да бъдат обяснени чрез разглеждане на различния ембрионален произход и начин на осификация на засегнатите скелетни елементи. разкриха вариации в изложените плодове по отношение на контролните, касаещи осификацията на черепните кости и ребрата на гръдната клетка, както и изместването на мекелийския хрущял. Колетата, изследвани след никнене на зъби, показват нормални фенотипове. Прави се заключението, че лекото излагане на радиация от мобилен телефон може да повлияе, макар и преходно, върху развитието на плода на мишката на ниво на осификация. Наблюдаваните вариации в развитието могат да бъдат обяснени чрез разглеждане на различния ембрионален произход и начин на осификация на засегнатите скелетни елементи. развитие на плода на мишката на ниво на осификация. Наблюдаваните вариации в развитието могат да бъдат обяснени чрез разглеждане на различния ембрионален произход и начин на осификация на засегнатите скелетни елементи. развитие на плода на мишката на ниво на осификация. Наблюдаваните вариации в развитието могат да бъдат обяснени чрез разглеждане на различния ембрионален произход и начин на осификация на засегнатите скелетни елементи.
Copyright © 2009 Elsevier Ireland Ltd. Всички права запазени.
Подобни статии
- Електромагнитното радиочестотно излъчване, излъчвано от GSM мобилни телефони, намалява точността на домашните монитори за кръвен глюкоза.J Biomed Phys Eng. 2014 г. 1 септември; 4 (3): 111-6. eCollection 2014 септември.PMID: 25505778 Безплатна статия от PMC.
- Излъчването на мобилен телефон по време на пубертетно развитие не оказва влияние върху тестологичната хистология при плъхове.Toxicol Ind Health. 2016 февруари; 32 (2): 328-36. doi: 10.1177 / 0748233713500820. Epub 2013 4 октомври.PMID: 24097363
- Когнитивно увреждане при плъхове след продължително излагане на излъчване на мобилен телефон GSM-900.Биоелектромагнетика. 2008 април; 29 (3): 219-32. doi: 10.1002 / bem.20386.PMID: 18044737
- Асоциация между използването на мобилен телефон и качеството на спермата: системен преглед и мета-анализ.Андрология. 2014 юли; 2 (4): 491-501. doi: 10.1111 / j.2047-2927.2014.00205.x. Epub 2014 3 април.PMID: 24700791 Преглед.
- Електромагнитни полета и ефекти върху здравето - епидемиологични изследвания на рак, заболявания на централната нервна система и сърдечни заболявания, свързани с аритмия.Scand J Work Environment Health. 2004; 30 Допълнение 1: 1-30.PMID: 15255560 Преглед.
Цитирано от 3 статии
- Има ли омега-3 защитен ефект върху надбъбречната жлеза на плъх, изложена на 900 MHz електромагнитни полета?J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 185-190. doi: 10.1016 / j.jmau.2017.08.003. Epub 2017 21 септември.PMID: 30023253 Безплатна статия от PMC.
- Ефекти от излагането на електромагнитни полета върху антиоксидантната защитна система.J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 167-176. doi: 10.1016 / j.jmau.2017.07.003. Epub 2017 2 август.PMID: 30023251 Безплатна статия от PMC. Преглед.
- Влиянието на електромагнитното излъчване, генерирано от мобилен телефон, върху костната система на плъхове.Biomed Res Int. 2015; 2015: 896019. doi: 10.1155 / 2015/896019. Epub 2015 1 февруари.PMID: 25705697 Безплатна статия от PMC.
LinkOut - повече ресурси
Източници на пълен текст
Дисбиндин модулира префронтални кортикални глутаматергични вериги и функция на работната памет при мишки. Jentsch, et al Neuropsychopharmacology 34, 2601–8 (2009).
Дисбиндин модулира префронтални кортикални глутаматергични вериги и функция на работната памет при мишки
Принадлежности- PMID: 19641486
- PMCID: PMC2762021
- DOI: 10.1038 / npp.2009.90
Резюме
Поведенческите генетични проучвания на хора свързват вариацията в гена DTNBP1 с шизофрения и нейните фенотипи на когнитивния дефицит. Протеинът, кодиран от DTNBP1, дисбиндин, се експресира в глутаматергичните неврони на предния мозък, в които той взаимодейства с протеини, участващи в трафика на везикули и екзоцитоза. За да се очертаят допълнително клетъчните, физиологичните и поведенческите фенотипове, свързани с намалена експресия на дисбиндин, проведохме проучвания върху мишки, носещи нулева мутация в гена dtnbp1. Мутантите на дисбиндин показват нарушения на пространствената работна памет в сравнение с контролите от див тип; хетерозиготни мишки показват междинни нива на когнитивна дисфункция. Дълбокослойните пирамидални неврони, записани в префронталната кора на мутантни мишки, показват намаляване на улеснението на сдвоения пулс, и предизвикани и миниатюрни възбуждащи постсинаптични течения, показващи разлика във функцията на пресинаптичните глутаматергични терминали, както и повишени прагове на скокове. Взети заедно, тези данни показват, че дисбиндинът мощно регулира възбудното предаване в префронталната кора, потенциално чрез предсинаптичен механизъм, и следователно модулира когнитивните функции в зависимост от този мозъчен регион, предоставяйки нови прозрения за молекулните механизми, в основата на кортикалната дисфункция при шизофрения
Изявление за конфликт на интереси
Разкриване / Конфликт на интереси Д-р Jentsch е получил обезщетение като консултант в Merck Research Laboratories. Другият автор (и) декларира, че с изключение на доходите, получени от основния ми работодател, не е получена финансова подкрепа или обезщетение от нито едно физическо или юридическо лице през последните три години за научни изследвания или професионално обслужване и няма лични финансови участия, които може да се възприеме като представляващ потенциален конфликт на интереси.
Фигури


Подобни статии
- Намалената експресия на дисбиндин медиира хипофункция на N-метил-D-аспартатния рецептор и нарушена работа на работната памет.Психиатрия на Biol. 2011 г. 1 януари; 69 (1): 28-34. doi: 10.1016 / j.biopsych.2010.09.012. Epub 2010 30 октомври.PMID: 21035792 Безплатна статия от PMC.
- Потенциални молекулярни механизми за намалено освобождаване на синаптичен глутамат при мишки с мутант на дисбиндин-1.Шизофр Рез. 2013 май; 146 (1-3): 254-63. doi: 10.1016 / j.schres.2013.01.037. Epub 2013 6 март.PMID: 23473812 Безплатна статия от PMC.
- Загубата на дисбиндин-1 компрометира NMDAR-зависимата синаптична пластичност и контекстното обуславяне на страха.Хипокампус. 2014 февруари; 24 (2): 204-13. doi: 10.1002 / hipo.22215. Epub 2013 12 ноември.PMID: 24446171 Безплатна статия от PMC.
- Допаминергичен контрол на работната памет и нейното значение за шизофренията: перспектива на динамиката на веригата.Неврология. 2006 г., 28 април; 139 (1): 153-71. doi: 10.1016 / j.neuroscience.2005.08.070. Epub 2005 1 дек.PMID: 16324800 Преглед.
- Пясъчната (sdy) мишка: мутант на дисбиндин-1, имащ отношение към изследванията на шизофренията.Prog Brain Res. 2009; 179: 87-94. doi: 10.1016 / S0079-6123 (09) 17910-4. Epub 2009 20 ноември.PMID: 20302821 Преглед.
Цитирано от 53 статии
- Гени за развитие и регулаторни протеини, домейни на когнитивно увреждане при психоза на шизофрения спектър и последици за откриването на антипсихотични лекарства: Примерът на изоформи на дисбиндин-1 и след това.Front Pharmacol. 2020 г., 29 януари; 10: 1638. doi: 10.3389 / fphar.2019.01638. eCollection 2019.PMID: 32063853 Безплатна статия от PMC. Преглед.
- Загубата на дисбиндин-1 засяга GABAergic предаването в PFC.Психофармакология (Берл). 2019 ноември; 236 (11): 3291-3300. doi: 10.1007 / s00213-019-05285-1. Epub 2019 14 юни.PMID: 31201475
- Свързване на двигатели с микротубули с органели, генерирани от AP-3 в аксони от калцион, член на семейството NEEP21.Mol Biol Cell. 2018 15 август; 29 (17): 2055-2068. doi: 10.1091 / mbc.E18-01-0007. Epub 2018 27 юни.PMID: 29949458 Безплатна статия от PMC.
- Дисбиндин-1 допринася за префронталната кортикална дендритна арборна патология при шизофрения.Шизофр Рез. 2018 ноември; 201: 270-277. doi: 10.1016 / j.schres.2018.04.042. Epub 2018 11 май.PMID: 29759351 Безплатна статия от PMC.
- Участие на дисбиндин-1 в етиологията на шизофренията.Int J Mol Sci. 2017 г., 22 септември; 18 (10): 2044. doi: 10.3390 / ijms18102044.PMID: 28937620 Безплатна статия от PMC. Преглед.
Препратки
- Arguello PA, Gogos JA. Моделиране на лудост при мишки: по едно парче. Неврон. 2006; 52 (1): 179–196. - PubMed
- Ballard TM, Woolley ML, Prinssen E, Huwyler J, Porter R, Spooren W. Ефектът на mGlu5 рецепторния антагонист MPEP при тестове за безпокойство и познание на гризачи: сравнение. Психофармакология (Berl) 2005; 179 (1): 218–229. - PubMed
- Bhardwaj SK, Baharnoori M, Sharif-Askari B, Kamath A, Williams S, Srivastava LK. Поведенческа характеристика на пясъчни мишки с дефицит на дисбиндин-1. Поведенчески изследвания на мозъка. 2008 - PubMed
- Bhardwaj SK, Baharnoori M, Sharif-Askari B, Kamath A, Williams S, Srivastava LK. Поведенческа характеристика на пясъчни мишки с дефицит на дисбиндин-1. Behav Brain Res. 2009; 197 (2): 435–441. - PubMed
- Bray NJ, Preece A, Williams NM, Moskvina V, Buckland PR, Owen MJ, et al. Хаплотипите на локуса на гена на дистробревин свързващ протеин 1 (DTNBP1) медиират риска от шизофрения чрез намалена експресия на DTNBP1. Човешка молекулярна генетика. 2005; 14 (14): 1947–1954. - PubMed
Видове публикации
Условия на MeSH
Вещества
Предоставяне на подкрепа
LinkOut - повече ресурси
Източници на пълен текст
Бази данни за молекулярна биология
Пътища за сигнализиране на стрес, които нарушават структурата и функцията на префронталната кора. Arnsten, AF National Review of Neuroscience 10, 410–22 (2009).Nat Rev Neurosci . Авторски ръкопис; на разположение в PMC 2010 20 юли.
Публикувано в окончателно редактирана форма като:Пътища за сигнализиране на стреса, които нарушават структурата и функцията на префронталната кора
Окончателната редактирана версия на тази статия на издателя е достъпна в Nat Rev NeurosciПрефронталната кора (PFC) интелигентно регулира нашите мисли, действия и емоции чрез обширни връзки с други мозъчни региони ( BOX 1 ). Той създава „ментална скица“ (за използване на фраза, измислена от Алън Баддели) чрез мрежи от неврони, които могат да поддържат информация при липса на стимулация от околната среда 1 . Невролози като Патриша Голдман-Ракич посочват този процес като работна памет: способността да се има предвид току-що настъпилото събитие или да се припомни информация от дългосрочно съхранение и да се използват тези представителни знания за регулиране на поведението, мисленето и емоция 2. PFC е в състояние да защити тези крехки изображения от намесата на външни или вътрешни разсейващи фактори и е от ключово значение за възпрепятстване на неподходящи действия и насърчаване на операции, свързани със задачата (така нареченото регулиране „отгоре надолу“) 3 - 6 . ПФК операции позволяват гъвкаво регулиране на поведението, за да можем правилно да отговори на променящата се среда - например, способността да се измести вниманието набор на нови измерения и с вземането на решение алтер като награда непредвидени смени 7 , 8 . PFC също наблюдава грешките, като ни дава представа, че сме грешни и трябва да променим стратегиите 9. Всички тези способности зависят от правилните връзки на невроналната мрежа на PFC, които са силно чувствителни към тяхната неврохимична среда.
Този преглед обсъжда как невромодулаторните промени, които се случват по време на стрес, бързо нарушават мрежовите връзки на PFC и значително увреждат функцията на PFC. Той се фокусира върху функциите на пространствената работна памет на PFC, тъй като схемите, физиологията и модулацията на дорзолатералните PFC неврони, които медиират работната памет, са най-добре характеризирани в този мозъчен регион. Прегледът първо описва как излагането на дори лек неконтролируем стрес може бързо да наруши функциите на PFC при хора и животни. След това се описват извънклетъчните и вътреклетъчните механизми, които допринасят за дефицита на PFC, и как хроничното излагане на стрес води до структурни промени в PFC. И накрая, той подчертава как генетичните и екологични промени в пътищата за сигнализиране на стреса са свързани с психични заболявания,
Остър стрес нарушава PFC функцията
Човешки изследвания
Някои от първите проучвания за въздействието на стреса върху познанието започват след Втората световна война, основавайки се на наблюдения, че пилотите, които са били висококвалифицирани по време на мирно време, често са разбивали самолетите си в стреса на битката поради психически грешки 10 . Започнаха изследвания за експериментално манипулиране на нивата на стрес, за да се види как това е променило производителността и когнитивните способности 11 . Много от тези ранни проучвания показват, че излагането на стрес влошава изпълнението на задачи, които изискват сложно, гъвкаво мислене, но че всъщност може да подобри изпълнението на по-прости и / или добре репетирани задачи 10 , 12 . Сега разбираме, че видовете задачи, които са били нарушени от стреса, са тези, които изискват PFC операции 13като има предвид, че вкоренените навици, които разчитат на вериги на базални ганглии, бяха спестени или подобрени 14 .
Тези ранни проучвания също посочват съществената роля на чувството за контрол на субекта върху стресора. Субектите, които се чувстваха контролирани от ситуацията (дори ако това беше илюзия), често не бяха нарушени от излагането на стрес, докато тези, които се чувстваха извън контрол, бяха нарушени 15 . Проучванията върху животни потвърдиха ключовата роля на контрола 16. Контролният фактор представлява особен проблем за съвременните изследвания на стреса при хора, тъй като етично на всички субекти трябва да бъде даден контрол върху тяхното положение и да могат да напуснат експеримента по всяко време. Независимо от това се появиха парадигми, които са ефективни за създаване на усещане за неконтролируем стрес в експериментална обстановка. Например, едно изследване с функционални образи е накарало дамите да гледат емоционално разстройващи филми и са открили доказателства за намалено активиране в PFC региони 17 . емоционалните разсейващи фактори могат по същия начин да намалят дорзолатералната PFC активност при субекти, изпълняващи задача за работна памет 18. Много проучвания на хормонални реакции на стрес са използвали задача за публично говорене като ефективен стрес (тест за социален стрес на Трир). Установено е, че този социален стресор уврежда когнитивната гъвкавост 19 и работната памет 20 , като и двете изискват PFC функция. Интересното е, че същата тази социална стресор всъщност подобрява класическа инсталация за отрицателни стимули, както и хипокампална пространствената памет 20 . Тези открития при хора съвпадат в голяма степен с проучвания върху животни, които показват, че остър лек стрес нарушава префронталната функция, но всъщност подобрява функционирането на амигдалата и хипокампуса (вж. По-долу). други проучвания са установили нарушена функция на PFC при субекти, използващи психични образи на собствените им преобладаващи преживявания 21 ,22 и при студенти по медицина, докато са учили за съветите 23 .
Намаляването на функционирането на PFC, което се случва по време на стрес, е от голямо значение за разбирането на човешкото психическо и физическо здраве. загубата на самоконтрол по време на излагане на стрес може да доведе до рецидив на редица дезадаптивни поведения, като пристрастяване към наркотици, пушене, пиене на алкохол и преяждане 22 . Продължителният стрес е основен рисков фактор за депресия 24 , 25 и излагането на травматичен стрес може да причини посттравматично стресово разстройство (ПТСР) 26 . Стресът може също така да изостри симптомите на шизофрения 27 , 28 и биполярно разстройство, например чрез превключване на пациентите от нормален период (евтимия) в маниакално състояние 29. Като се имат предвид тези мощни ефекти върху човешкото здраве, важно е да има животински модели на реакцията на стрес, които да ни помогнат да разберем механизмите, които ни правят уязвими.
Проучвания върху животни
Ранните проучвания на въздействието на стреса върху производителността при гризачи използват така наречената парадигма на научената безпомощност. Тези проучвания са сред първите, които показват, че неконтролируемият стрес (т.е. неизбежен шок), но неконтролируемият стрес, нарушават изпълнението на задача Y-лабиринт поради дефицити в селективното внимание 16 . По-скорошни проучвания, използващи тази парадигма, показват, че когато животното възприема себе си като контролиращо, PFC е ключов за потискане на реакциите на стрес, предизвикани в мозъчния ствол 30 . Всъщност PFC и хипокампусът са двете мозъчни структури, които са позиционирани да инхибират реакцията на глюкокортикоиден стрес 31 .
Няколко проучвания са използвали задачи, които изрично разчитат на функцията на PFC, за да изследват ефектите на лекия стрес върху когнитивните резултати. Тези изследвания се използват пространствени задачи работна памет в плъхове и маймуни и е установено, че съвсем леко остро стресово нарушена точността на отговор и често предизвиква perseverative модел на отговор, който е в съответствие с PFC дисфункция 32 - 34 . Например бе установено, че стресът от бял шум, който уврежда когнитивните способности на хората, също нарушава пространствената работна памет при маймуните 33. Обратно, изпълнението на контролни задачи със сходни двигателни и мотивационни изисквания, но липсата на необходимост от регулиране на PFC не е променено от лекото излагане на стрес. По същия начин плъховете, изложени на остри стресови фактори, са били увредени при задача за пространствено забавено редуване, която изисква медиална PFC функция, но не са били увредени при независима от PFC задача за пространствена дискриминация в същия лабиринт 32 .
За разлика от това, острото излагане на лек стрес не оказва влияние или може действително да подобри функциите за консолидиране на паметта на хипокампуса и амигдалата 35 , 36 . (Обърнете внимание, че вентромедиалният PFC регулира реакциите на страх, медиирани от амигдалата; вижте REF. 37 за скорошен преглед.) По-тежките остри стресови фактори нарушават функциите на хипокампата 38, но продължават да укрепват емоционалните асоциации и двигателните навици, осъществявани от амигдалата и стриатума , съответно 14 , 39 . По този начин, остър неконтролируем стрес нарушава опосредстваните от PFC когнитивни функции при хора и животни и превключва контрола на поведението и емоциите към по-примитивни мозъчни вериги.
Катехоламините медиират ефектите на остър стрес
Как неврохимичните промени, предизвикани от излагане на остър стрес, влошават функцията на PFC? Способностите на работната памет на PFC в решаваща степен зависят от точната активност на взаимосвързаните невронни мрежи. Микросхемата, която поддържа пространствената работна памет при приматите, е привлекателен модел за изследване, тъй като анатомията, физиологията и невромодулацията на тези мрежи са добре характеризирани благодарение на пионерската работа на Goldman-Rakic и колеги 40. Този преглед се фокусира върху катехоламиновата модулация на вериги за пространствена работна памет, тъй като в момента това е арена, където най-добре разбираме как подобните на стрес промени в химическата среда променят моделите на невронална реакция в префронталните мрежи на приматите, които активно участват в когнитивна операция. Надяваме се, че бъдещите проучвания ще разширят този подход върху други области на кората, допълнителни когнитивни операции и други ключови невромодулатори.
Схемите, които са в основата на пространствената работна памет, са картографирани с много подробности. В кората на приматите силно обработената визуална пространствена информация се подава напред от корите на теменната асоциация към основната сулкална кора в дорзолатералния PFC 40 . Тази област е от решаващо значение за пространствената работна памет, тъй като лезиите в тази област, но не и в околната тъкан, водят до дълбоки и постоянни дефицити по задачите за пространствена работна памет 40 . Микросхемите в PFC могат да поддържат представяне на визуално пространствена информация при липса на стимулация на околната среда - например по време на периода на закъснение в работна памет. Funahashi et al . 41 разработи задача за сондиране на веригата за пространствена работна памет, наречена задача за окуломоторен забавен отговор (ODR) (Фиг. 1). В тази задача маймуната трябва да запомни точно пространствено местоположение за период на забавяне, преди да премести очите си на запомненото място, за да получи награда за сок (Фиг. 1а). всяка сесия се състои от стотици опити, като правилната пространствена позиция се променя между опитите. По този начин съдържанието на работната памет трябва постоянно да се актуализира. От PFC се записват единични единици (Фиг. 1б), докато маймуната изпълнява задачата за ODR. В този регион много неврони показват силно настроена, постоянна активност по време на периода на забавяне; тази дейност представлява пространствено положение при липса на стимулация на околната среда (например вжФиг. 1в). Счита се, че персистирането на невронната активност по време на периода на забавяне възниква от повтарящо се възбуждане между PFC пирамидални клетки със споделени входове от теменната кора - т.е. между пирамидални клетки със сходни пространствени характеристики (Фиг. 1г). Тези възбуждащи повтарящи се връзки вероятно включват NMDA ( N -метил- d- аспартат) и AMPA (α-амино-3-хидрокси-5-метил-4-изоксазол пропионова киселина) рецепторни синапси върху дендритни бодли. За разлика от това, високата степен на пространствена настройка произтича от активността на локалните инхибиторни интернейрони (например GABA (γ-аминомаслена киселина) -ергични клетки на кошницата и полилейни клетки (Фиг. 1г)), които осигуряват странично инхибиране. По този начин, когато са активни пирамидални клетки, представляващи например пространствени позиции ∼90 ° от централна референтна точка, те се възбуждат взаимно, за да създадат постоянна активност и в същото време активират GABAergic интернейрони, които потискат активността на пирамидалните клетки, представляващи други посоки като клетките, посветени на 45 °). В този пример 90 ° ще се нарече „предпочитана посока“ на възбудените неврони, а други пространствени позиции ще се наричат „нежелани посоки“ за тези неврони. Въпреки че глутаматът и GABA са основата на това пространствено настроено, мнемонично изпичане, силата на постоянното изпичане и степента на пространствена настройка също са силно повлияни от катехоламините норадреналин и допамин.
Пространствени мрежи за работеща памет в дорзолатералната префронтална кораa | Задачата на окуломоторната забавена реакция (ODR) е задача за пространствена работна памет, която се използва за изследване на физиологичните профили на невроните на префронталната кора (PFC). Субектът трябва да запомни пространствената позиция на най-новата реплика за период на забавяне от няколко секунди и след това да посочи тази позиция със сакада до запаметеното място. b | Регионът на дорсолатералния PFC на маймуната (зона на Уокър 46), където невроните показват постоянна, пространствено настроена стрелба по време на периода на закъснение в задачата за ODR. ° С| Пример за PFC неврон, който показва постоянна стрелба по време на периода на закъснение, ако репликата е възникнала при 90 ° - „предпочитаната посока“ за този неврон (ляв среден график) - но не и ако репликата се появява в „непредпочитан“ посока (други парцели). Растерите показват изстрелването на неврона в продължение на седем опита във всяка пространствена позиция. г | Схематичен чертеж на PFC микросхемите, които са в основата на пространствената работна памет, както е описано от Goldman-Rakic 40. Пирамидалните клетки от слой III получават силно обработена пространствена информация (представена от зелените линии) от корите на теменната асоциация. Пирамидалните клетки с подобни пространствени характеристики участват в повтарящо се възбуждане, за да поддържат постоянна активност през периода на забавяне (имайте предвид, че субклетъчната локализация на тези възбуждащи връзки понастоящем не е известна; те могат да бъдат върху апикалните и / или базалните дендрити). GABA (γ-аминомаслена киселина) -ергични интернейрони, като клетките на кошницата (B) и полилеите (C), помагат за пространствената настройка на невроните чрез странично инхибиране. Входните данни от кръстосани микросхеми (неврони с различни настройки за настройка) са показани в сиво. AS, дъговидна бразда; PS, основна бразда. Части а – в са модифицирани с разрешение от REF. 60 © (2007) Cell Press.
Норадреналиновите и допаминовите неврони в мозъчния ствол променят скоростта си на стрелба според състоянието ни на възбуда и според значението на събитията в околната среда. Норадреналин неврони в coeruleus процесуалната са тихи по време на бързия сън движение на очите и имат ниска тоник стрелба по време на сън с бавни вълни, умерена тоник изпичане и ясно изразен стадиален изпичане на съответните стимули по време на които не са подчерта, събуждане, и високо тоник стрелба с нарушена регулация стадиален стрелба по време на стрес 42 . Те стрелят към съответните стимули по време на будно събуждане, но могат да стрелят към неподходящи стимули (дистрактори) по време на умора или стрес 42 . Допаминовите неврони не са били проследявани по методичен начин по отношение на състоянията на възбуда, но като цяло увеличеното фазово изстрелване е свързано с прогнозиране на наградата 43. Неотдавнашни проучвания обаче предполагат, че подгрупа от допаминови неврони на средния мозък увеличава стрелбата си до аверсивни стимули 44 и е възможно те да са в основата на промените във функцията на PFC, които се появяват по време на стрес. Важно е, че освобождаването на допамин и норадреналин се увеличава в PFC по време на излагане на остър стрес 45 , 46 . Всъщност дори много лекият стрес увеличава освобождаването на допамин в PFC 47 .
Въпреки че инервацията на норадреналин и допамин на PFC е доста деликатна 48 , тя е изключително мощна. Норадреналинът и допаминът имат „обърнато U-образно влияние върху работната памет, при което или твърде малко, или твърде много норадреналин и / или допамин нарушават PFC функцията (Фиг. 2). Норадреналинът и допаминът осигуряват съществени възбуждащи влияния, които поставят кората в будно състояние, в което невроните могат да обработват и обменят информация 49 , 50 . Те обаче имат и допълнителни модулаторни влияния, които силно влияят върху силата на PFC връзките, когато мрежите участват в операции с работеща памет.
Катехоламинът влияе върху физиологията и функцията на префронталната кораКакто норадреналинът (NA, част а ), така и допаминът (DA, част б ) имат „обърнато U-образно“ влияние върху физиологията и познанието на префронталната кора (PFC), при което твърде малко или твърде много от невротрансмитера нарушава функцията на PFC. В задача за забавен отговор на окуломотор, с оптимални нива на освобождаване на NA или DA при аларма, ненатоварени условия (в горната част на кривите), PFC невроните се задействат (както е показано в зелените следи) по време на периода на закъснение след сигнали за предпочитани, но не не-предпочитани упътвания. NA засилва свързаното със забавяне стрелба в отговор на сигнали в предпочитани посоки чрез стимулиране на α 2A-рецептори (увеличаване на „сигнала“), докато DA отслабва свързаното със забавяне стрелба в отговор на сигнали в непредпочитани посоки чрез стимулиране на D1 рецепторите (намаляване на „шума“). Прилагането на подходящи концентрации на α2A- рецепторен агонист гуанфацин или D1 рецепторен агонист SKF81297 също има този ефект. С високи нива на освобождаване на NA по време на стрес (дясната страна на кривата), NA ангажира α 1 -рецепторите с по-нисък афинитет и така намалява стрелбата на невроните. По същия начин, прекомерната стимулация на D1 рецепторите по време на стрес потиска стрелбата на клетките. Приложение на α 1 -рецепторен агонист фенилефрин или висока концентрация на SKF81297може да имитира ефектите на високи нива на NA и DA, съответно. Във всяка графика пунктираните линии показват прехода между (отляво надясно) периода на фиксиране, периода на реплика, периода на забавяне и периода на реакция на окуломотора.
Обърнатата U-образна връзка между нивата на норадреналин и работната памет е особено интересна, тъй като норадреналинът ангажира различни видове рецептори с различни нива на освобождаване. Норадреналинът има най-висок афинитет към α 2- адренергичните рецептори и по-нисък афинитет към α 1 - и β-адренергичните рецептори 51 . Както е показано вФиг. 2, нивата на норадреналин, които се освобождават по време на тревога, нестресирано събуждане, оптимизират работната памет чрез включване на α2A- рецептори 52 , 53 , докато високите нива на норадреналин, които се отделят по време на стрес, влошават функцията на PFC чрез стимулиране на по-нисък афинитет α 1 - рецептори 54 и β 1 -рецептори 55 . По този начин или изчерпването на норадреналин 52, или блокадата на α 2A -рецептори в PFC 53 уврежда работната памет, докато стимулирането на постсинаптичните α 2A -рецептори в PFC подобрява производителността на работната памет 52 , 56 -58 . Тези ефекти могат да се видят на клетъчно ниво също: неадекватна α 2А рецепторен стимулация намалява PFC стрелба, докато α 2А рецепторен стимулация в ПФК подобрява задействане на невроните в периода закъснение за предпочитана посока невроните "в задачата за ОРС 59 , 60 (Фиг. 2а). За разлика от това, високите нива на стимулация на 1- рецептора в PFC бързо потискат изстрела на невроните и влошават работата на пространствената работна памет 61 , 62 . По същия начин, предизвикани от стрес когнитивни дефицити могат да бъдат предотвратени чрез прилагане на а 1 рецепторни антагонисти в PFC или системно 54 . Тези открития са в съответствие с клинични изследвания показват, че алфа 1 рецепторен антагонист празосин е полезно при лечение на посттравматично стресово разстройство 63 , 64 . Проучванията при хора също са установили, че антагонист на β-рецептор може да попречи на индуцирана от стрес увреждане на познавателната гъвкавост 19. По този начин има добро съгласие между проучванията върху животни и хора.
Допаминът упражнява своето обърнато U-образно влияние върху способностите на работната памет чрез действия в семейството на D1 рецепторите (тъй като настоящите лекарства не могат да дисоциират D1 и D5 рецепторите, съответният им принос не се разбира). И блокадата, и прекомерното стимулиране на D1 рецепторите увреждат пространствената работна памет 65 - 67 . Дефицитите на работната памет по време на излагане на стрес включват прекомерна стимулация на D1 рецептора, тъй като те се предотвратяват чрез приложението на D1 антагонист 32 . Обърнатата U-образна връзка може да се наблюдава и на клетъчно ниво (Фиг. 2б), където оптималното ниво на D1 рецепторна стимулация подобрява пространствената настройка чрез потискане на свързаното със забавяне стрелба към нежеланите посоки на невроните в задачата за ODR 68 . Така, допамин и норадреналин имат допълнителни функции: α 2А рецептор стимулация подобрява мрежа изпичане на споделени ресурси (това означава, че увеличава "сигнал" (Ref. 60 )), докато D1 рецептор за стимулиране на невронна вае изпичане чрез намаляване на изпичане на не- предпочитани входове (т.е. намалява „шума“ (РЕФ. 68 )). Въпреки това, при по-високи нива на D1 рецепторна стимулация, стрелбата е потисната за всички посоки и невронът губи както пространствената си настройка, така и нивото на отзивчивост 68 (Фиг. 2б). По този начин, високите нива на катехоламини по време на стрес намаляват както постоянното изстрелване, така и настройката на PFC невроните. Вероятно D2 семейството на допаминовите рецептори също допринася за реакцията на стрес. прекомерното стимулиране на D2 рецепторите нарушава функцията на PFC на работната памет при животни 69 и хора 70 и е свързано с повишено стрелба при маймуни 71, свързано с отговора, и повишени сигнали, зависими от нивото на кислород в кръвта в кръвта при проучвания при изображения на хора 70 . Обърнатата u-образна връзка между нивата на допамин и функционирането на PFC е наблюдавана при хора както във фармакологични проучвания 72, така и по отношение на катехол- О- метилтрансферазата ( COMT) генотип 73 : заместването на метионин в протеина COMT, което е резултат от един от полиморфизмите на COMT, намалява способността на COMT да катаболизира допамина и увеличава уязвимостта към предизвикани от стрес или стимуланти нарушения на работната памет при хора 74 и при мишки 75 .
Трябва да се отбележи, че стрес-индуцирана увреждания в работната памет могат да бъдат спасени чрез блокиране или D1 или а 1 рецептори в PFC. Тези открития са в съответствие със синергичните взаимодействия между ефектите на допамин и норадреналин. Катехоламинните действия могат също така да синергизират с глюкокортикоидните ефекти в PFC. Стресът увеличава освобождаването на глюкокортикоиди, както и това на катехоламини, и проучвания както при животни 76, така и при хора 77са показали, че високите нива на глюкокортикоиди сами по себе си могат да увредят работната памет. Тези ефекти на глюкокортикоидите са бързи и вероятно не се проявяват чрез традиционните геномни механизми. Възможно е глюкокортикоидите да преувеличават катехоламиновите действия в PFC, като блокират екстраневроналните катехоламинови транспортери върху глията, които изчистват екстрасинаптичното пространство от катехоламини 78 . Подобна синергична връзка между глюкокортикоидите и котешките ехоламини вече се наблюдава в amyg dala, където глюкокортикоидите и действията на 1 - и β-рецепторите подсилват консолидацията на паметта 79 , 80 .
Амигдалата играе ключова роля в промяната на неврохимичната среда на PFC по време на стрес. лезиите на амигдалата предотвратяват увеличаването на освобождаването на допамин и норадреналин, което се случва в PFC в отговор на психологически стрес 81 . Както е показано в КЛЕТКА 1 , при стресови условия проекциите на амигдала към мозъчния ствол и към хипоталамуса стимулират освобождаването на катехоламини и глюкокортикоиди през невраксиса. Тези високи нива на катехоламини и глюкокортикоиди укрепват функциите на амигдала, като кондициониране на страха и консолидиране на емоционално значима информация 79 , 80, но отслабват функционирането на PFC; по този начин те изместват оркестрацията на мозъчните реакции от PFC към амигдалата. Високите нива на катехоламини също засилват функционирането на други подкоркови мозъчни региони и задните кортикални области. Например, високите нива на норадреналин, действащи при α 1 - и β-рецепторите, подобряват процесите на консолидация на паметта в хипо-кампуса 82 , 83 и увеличават съотношението сигнал / шум в първичните сензорни корти 84 - 86 . По-високите нива на освобождаване на допамин също насърчават формирането на навици в базалните ганглии 87. Възможно е високите нива на освобождаване на допамин в схемите на кортикобазални ганглии по време на стрес да служат за улавяне на каквото и да е успешно поведение, спасило субектите от опасност, и да закрепят този модел като навик. Но същото това еволюционно решение може да направи хората уязвими към дезадаптивно поведение (например моторни тикове, наркомании и слухови халюцинации), когато неподходящ поведенчески модел е „уловен“ от високите нива на допамин. Взети заедно, доказателствата сочат, че високите нива на освобождаване на катехоламин, инициирани от амигдала по време на стрес, превключват мозъка от внимателна, отразяваща регулация от PFC към по-бърза рефлексивна регулация от амигдала и други подкоркови структури ( BOX 1). Тези механизми могат да спасят живота ни, когато сме в опасност и трябва да реагираме бързо, но те могат да навредят, когато трябва да направим избор, който изисква внимателен анализ и инхибиторен контрол.
Вътреклетъчни сигнални пътища
Вътреклетъчните механизми, които причиняват загубата на PFC функция на работната памет по време на остър стрес, вече започват да се разбират и те дават важни улики за разбиране на генетичната основа и лечение на психични заболявания. Обобщение на тези сигнални пътища и техните взаимодействия може да се види вФиг. 3.
Вътреклетъчни сигнални пътища, които нарушават функциите на префронталната кора на работната памет по време на стресВътреклетъчните сигнални пътища, активирани от излагане на стрес, имат обратни взаимодействия, които бързо увреждат когнитивната функция на префронталната кора (PFC). Високите нива на допамин (DA) D1 рецепторна стимулация (и вероятно норадреналин (NA) β 1 рецепторен стимулация, както и) активиране на аденилил циклази (ACS) до получаване на цикличен AMP. cAMP отваря активирани от хиперполяризация циклични нуклеотидни катионни канали (HCN канали) върху дендритни шипове, за да произведе h ток (I h ), който отслабва мрежовите входове и намалява закъснението. Високите нива на NA също стимулират α 1 -рецептори, които активират сигнализирането на фосфатидилинозитол бифосфат (PIP2) -протеин киназа C (PKC). Последващ инозитол-1,4,5-трисфосфат (InsP 3 ), медииран CaДоказано е, че освобождаването 2+ намалява задействането на PFC клетки чрез отваряне на SK канали, което води до ток (I SK ), и е показано, че поддържа изстрелване през деполяризиращия ток I CAN . I CAN може да бъде намален с PKC и cAMP, така че потискащите ефекти на I SK вероятно преобладават при условия на стрес. Двата сигнални пътя си взаимодействат, за да засилят действията си (пунктирани стрелки); например, сАМР може да потенцира InSP 3 -medicated Са 2+ освобождаване чрез протеин киназа-медиираното фосфорилиране на инспектор 3 рецептори. И обратно, Ca 2+може да активира много изоформи на AC, за да генерира повече cAMP. Глюкокортикоидите също могат да активират тези пътища (виж основния текст). Ензимите, които обикновено осигуряват молекулярните спирачки по тези сигнални пътища за стрес (нарушени при шизофрения 1 (DISC1), регулатор на сигнализирането на G-протеин 4 (RGS4) и DAG киназа (DGK)), често са генетично променени в семейства със сериозни психични заболявания, поради което увеличаване на податливостта на излагане на стрес. Обърнете внимание, че показано е също така, че сигналните пътища на cAMP и PKC се активират в амигдалата по време на стрес, където засилват дългосрочната консолидация на паметта и кондиционирането на страха. DAG, диациглицерол; PDE4B, фосфодиестераза 4В.
Високите нива на стимулация на α 1 -адренергичния рецептор по време на стрес влошават функцията на работната памет на PFC чрез активиране на вътреклетъчните сигнални пътища на фосфатидилинозитол-протеин киназа С (PKC). При гризачи дефицитите работната памет, които са причинени от фармакологичен стрес, студен воден стрес или приложение на алфа 1 -агонисти може да бъде обърната чрез вливане на РКС инхибитори директно в PFC 61 , 88 . И обратно, предизвиканите от стрес увреждания на работната памет могат да бъдат имитирани чрез вливане на PKC активатор в PFC 61 . Подобни ефекти се наблюдават на клетъчно ниво: потискане на стрелба на PFC неврон, предизвикана от α 1-рецепторната стимулация може да бъде обърната чрез йонофореза на РКС инхибитор 61 . Записите in vitro от PFC срезове показват, че рамото на инозитол-1,4,5-трис-фосфат (InsP 3 ) -Ca 2+ от фосфатидилинозитол също е важно 89 . Както е показано вФиг. 4g, активирането на фосфатидилинозитол сигнализиране освобождава InsP 3 , който може да освободи Ca 2+ от вътреклетъчните запаси в дендритите. С адекватно поколение InsP 3 се задействат вълни от освобождаване на Ca 2+ , които могат да се придвижват към сомата и да потискат изстрела на невроните чрез отваряне на канали с малка проводимост, активирани Ca 2+ / K + (SK канали) върху сомата, което води до ток (I SK ) 89 . В допълнение, освобождаването на Ca 2+ улеснява PKC активността, като го кара да се транслокира от цитозола към мембраната, където се активира от диацилглицерол (DAG) (Фиг. 3). Вътреклетъчното освобождаване на Ca 2+ също активира деполяризиращ ток I CAN , който възстановява невроналното изстрелване 89 (Фиг. 3). Въпреки това, I CAN се потиска от високи нива на PKC и циклична активност на AMP и по този начин може да не възстанови нормалното стрелба при условия на стрес, когато активността на PKC и cAMP е висока.
Работещ модел, показващ как пирамидалната клетъчна активност на префронталната кора може да се регулира при оптимални условия спрямо стресa | Ефективността на мрежовите входове върху дендритните шипове на апикалните и / или базалните дендрити се регулира динамично чрез сигнализиране на цикличен AMP – хиперполяризационен цикличен нуклеотид-катионен канал (HCN канал). При оптимални условия подходящите мрежови връзки се укрепват от α 2A-рецепторно инхибиране на сигнализиране на cAMP – HCN канал, докато неподходящите мрежови връзки са отслабени чрез активиране на допамин D1 рецептора на сигнализиране на cAMP – HCN канал. Малкият обем на отделението на гръбначния стълб вероятно позволява много локализирана регулация на близките HCN канали. Полученият в резултат специфичен модел на мрежова свързаност позволява точно представяне на пространственото положение на репликата в задачата за забавена реакция на окуломотора и широчината на свързаността може да бъде динамично регулирана въз основа на текущите когнитивни изисквания. b | HCN каналите на плазмената мембрана на дендритните валове вероятно регулират възбудимостта. Те може да не са отворени, когато дендритът е деполяризиран от голям брой възбуждащи мрежови връзки. ° С| При оптимални условия тялото на клетката вероятно получава мрежови сигнали и съответно се задейства. г | При оптимални условия PFC показва силно моделирана активност, при което специфични подмножества на невроните са активни, за да представят точното пространствено положение на стимула. Това е обозначено на фигурата, на която е активна само микросхемата, представляваща 90 °. Имайте предвид, че интернейроните не са показани в тази диаграма за по-голяма яснота. Мрежовите входове са показани на апикалните дендрити, но всъщност може да са на базалните дендрити. Червените кръгове представляват α 2A -рецепторно укрепване на мрежовите входове; сивите кръгове представляват D1 рецептор-cAMP отслабване на мрежова връзка. д| При условия на стрес високите нива на cAMP отварят HCN канали в целия дендрит, отслабвайки всички мрежови връзки. е | При загуба на мрежови възбуждащи входове дендритът може да хиперполяризира и HCN каналите на дендритните шахти могат да се отворят, за да помогнат за поддържането на дендритната възбудимост. Отварянето на HCN канал също би намалило ефективността на входовете към дисталните части на дендрита. g | Високите нива на сигнализиране на фосфатидилинозитол по време на стрес биха увеличили медиираните от инозитол-1,4,5-трисфосфат (InsP 3 ) вълни на вътреклетъчно освобождаване на Ca 2+ . Когато Ca 2+ вълните нахлуят в сомата, те отварят малка проводимост, Ca 2+ активиран K +канали (SK канали) и намаляват стрелбата на клетките. h | При стресиращи условия мрежовите връзки са отслабени (сиви кръгове) и в резултат на това задействането на мрежата е намалено и без шарки. По този начин невроните на префронталната кора не могат да представят точно информацията в работната памет. Нехарактерната, генерализирана дейност може да изпълнява друга функция - например стимулиране на стресовите пътища в мозъчния ствол.
Високите нива на катехоламини по време на стрес също могат да нарушат функционирането на PFC чрез прекомерна cAMP сигнализация. Вътреклетъчната сигнална каскада cAMP има мощни ефекти върху мрежовата активност на работната памет и когнитивните функции. cAMP често действа чрез сигнализиране на протеин киназа А (PKA) и е вероятно много пластични процеси в PFC да изискват PKA сигнализация, точно както пластичните процеси в други мозъчни области правят 90 . Например, консолидацията на паметта при дълги закъснения (~ 30 минути или повече) изисква префронтално-хипокампални взаимодействия, които се възползват от cAMP активирането на PKA. Въпреки това, сАМР може да действа и на две други цели, а именно обмен на протеин, активиран от сАМР ( EPAC ; известен също като RAPGEF3) 91и активирани с хиперполяризация циклични нуклеотидни катионни канали (HCN канали) 92 . Физиологичните данни показват, че cAMP може да има особено мощни ефекти върху работещите мрежи памет чрез действия върху HCN канали. cAMP увеличава вероятността HCN каналите да се отворят 93 и да позволят преминаване както на Na +, така и на K + (това се нарича също h h или I h ). HCN каналите са локализирани както върху дендритните валове, така и върху дендритните шипове на PFC пирамидални клетки 60 (Фиг. 4). HCN каналите на дендритните валове вероятно имат традиционната роля да помагат за деполяризация на неврона, когато клетката е станала хиперполяризирана. Всъщност обширната блокада на HCN канали с високи дози ZD7288 предизвиква загуба на клетъчно изстрелване, в съответствие със загубата на това деполяризиращо влияние 60 . Въпреки това, HCN каналите на шипове изглежда имат по-изящна роля, като пренасочват мрежови входове към шипове. HCN каналите са локализирани до α2A- адренергичните рецептори 60или допаминови D1 рецептори (C. Paspalas и AFTA, непубликувани наблюдения) върху бодли в близост до асиметрични (вероятно възбуждащи) синапси. По този начин, катехоламиновите рецептори могат да регулират нивата на сАМР в затворено клетъчно отделение и така да определят състоянието на HCN каналите (отворени или затворени) близо до синаптичните входове на гръбначния стълб (Фиг. 4а). Когато HCN каналите се отворят в присъствието на cAMP, те шунтират близките входове. Последните данни показват, че това шунтиране възниква от отварянето на K + канали (Kv7 канали), което води до хиперполяризиращ М ток 94 . Високите нива на cAMP могат да потенцират тези действия чрез PKA модулация на Kv7 канали 95 , като по този начин се произвежда координирано cAMP шунтиране на мрежови входове.
При оптимални условия на възбуда (предупреждение, но без стрес), отварянето на HCN и Kv7 канал ще бъде строго регулирано. Физиологичните и поведенчески данни показват, че при такива условия α2A- рецепторна стимулация укрепва мрежовите входове от неврони със споделени пространствени характеристики чрез инхибиране на cAMP – HCN (и вероятно cAMP – PKA – Kv7) сигнализиране 60 . Например, блокиране на HCN каналите с ниски дози от ZD7288 обръща ефектите на α 2А рецептор блокада и възстановява нормалната активност на мрежата 60 . По подобен начин, вливането на ниски дози ZD7288 или нокдаун на HCN1 канали в PFC на плъхове подобрява производителността на работната памет 60. И обратно, ние предлагаме, че в задача за ODR, D1 рецепторите потискат невронните реакции към непредпочитани посоки чрез увеличаване на производството на сАМР 68 и отваряне на HCN канали на шипове, които получават мрежови входове от клетки, настроени към тези други посокиФиг. 4а). По този начин, при оптимални условия производството на сАМР ще бъде строго контролирано и ограничено до специфична подгрупа от бодли, получаващи вход от неврони, които не са от значение за текущите изисквания на задачата. при тези условия HCN каналите върху дендритите ще бъдат затворени и изстрелването на PFC клетки ще бъде силно моделирано (Фиг. 4г).
За разлика от това, в условия на неконтролируем стрес има прекомерно производство на сАМР и увреждане на работната памет. Нарушението в работната памет, предизвикано от излагане на стрес или прекомерна стимулация на D1 рецептор, може да бъде предотвратено чрез блокиране на cAMP активност или HCN канали в PFC 60 , 68 (AFTA, непубликувани наблюдения) или може да бъде имитирано чрез администриране на cAMP аналогов Sp-cAMPS 96 . Подобни резултати се наблюдават на клетъчно ниво, при което Sp-cAMPS, високи дози D1 агонист или инхибитор на фосфодиестераза потискат невроналното стрелба при маймуни, изпълняващи задача с работна памет, докато инхибирането на cAMP или нежната блокада на HCN каналите възстановява нормалните модели на стрелба 60 , 68. Тези открития са схематично илюстрирани вФиг. 4д, е, които показват, че високите нива на сАМР по време на стрес отварят HCN канали през дендрита, отслабвайки мрежовите входове от всички посоки (Фиг. 4д). загубата на възбуждащи мрежови входове на шипове би хиперполяризирала клетката, което би могло да отвори HCN канали на дендритни шахти, за да помогне за поддържането на известно изстрелване на клетката (Фиг. 4f). при тези условия клетката би показала силно намалена и нестандартна активност, както е схематично изобразена вФИГУРИ 2,U, 4ч4ч.
Пътищата за сигнализиране на стреса имат няколко взаимодействия, които могат допълнително да допринесат за бързите и драматични промени в мозъчните реакции на остър стрес (Фиг. 3). Например, cAMP-PKA сигнализирането може да стимулира фосфорилирането на InsP 3 рецепторите, което насърчава освобождаването на Ca 2+, медиирано от InsP 3 97 . И обратно, вътреклетъчното освобождаване на Са 2+ от повишеното сигнализиране за фосфатидилинозитол може да активира чувствителни към Са 2+ аденил циклази, за да увеличи сигнализирането на cAMP 98 (Фиг. 3). Сигнализирането както на cAMP 99, така и на PKC 100 намалява деполяризиращия ток I CAN , което допълнително би отслабило постоянното стрелба 89 , 101 , 102 . Освобождаването на глюкокортикоиди по време на стрес може допълнително да потенцира тези пътища чрез негеномно активиране на PKC сигнализиране 103 (въпреки че не е известно конкретно дали глюкокортикоидите имат този ефект в PFC). Тези синергични взаимодействия биха били необходими за бързо превключване на контрола на поведението от PFC към по-рефлексивни подкоркови пътища при наличие на остра опасност.
Ефекти от хроничен стрес върху PFC
В сравнение с излагането на остър стрес, излагането на хроничен стрес води до по-големи промени в прелимбичния PFC на плъхове, включително архитектурни промени. Невроните от слой II / III, които образуват PFC мрежи, губят дендритен материал - т.е. дължината на дендрита, разклоняването и плътността на гръбначния стълб се намаляват от хроничен стрес 104 , 105 . Загубата на гръбначен стълб е особено очевидна на дистални места (∼200 µm от сомата), а гъбовидните бодли, които вероятно участват в установени синаптични връзки, изглеждат особено уязвими 106 . Тези дендритни промени в PFC на плъхове са свързани с подчертана дисфункция на PFC при смяна на вниманието 107и работна памет (AB Hains и AFTA, непубликувани наблюдения). Хроничният стрес също така нарушава пластичната връзка между PFC и хипокампуса, която е необходима за гъвкаво консолидиране на паметта 108 . Дендритните промени в PFC постепенно се обръщат, когато стресът намалее 109 .
Както при острия стрес, PFC изглежда е особено чувствителен към архитектурни промени, предизвикани от хроничен стрес, в сравнение с други мозъчни региони. Докато структурните промени в хипокампуса изискват няколко седмици излагане на стрес 35 , дендритите в PFC започват да се променят само след една седмица на стрес 110 или дори дори еднократна експозиция 111 . За разлика от PFC и хипокампуса, дендритите в амигдалата се разширяват в отговор на излагане на хроничен стрес 112. По този начин хроничният стрес отслабва структурите, които осигуряват отрицателна обратна връзка за реакцията на стрес и укрепва структурите, които насърчават реакцията на стрес. Интересното е, че последните данни показват, че невроните в инфралимбичния PFC на плъхове, които се проектират към амигдалата, не губят своите дендрити в отговор на стрес, подчертавайки отчетливия отговор на веригите на амигдала 113 .
Сигналните механизми, които са в основата на дендритните промени в PFC, едва започват да се изучават. Хроничният стрес променя пътищата на катехоламин, увеличавайки норадренергичната инервация на PFC 114 (въпреки че допаминът се изчерпва при тежък хроничен стрес 115 ). Повишеният норадреналин може да доведе до по-високи нива на PKC и cAMP сигнализиране. Установихме, че повишената PKC сигнализация допринася за загубата на гръбначния стълб, тъй като ежедневното лечение с PKC инхибитор защитава плътността на гръбначния стълб и работната памет на PFC от вредните ефекти на хроничния стрес (AB Hains и AFTA, непубликувани наблюдения). Индуцираните от стреса промени в дендритната морфология могат да бъдат имитирани чрез хронично приложение на високи дози глюкокортикоиди 116 , 117, което може да включва екситотоксичност или оксидативен стрес 118 . Хроничната експозиция на глюкокортикоиди също намалява нивата на невротрофичния фактор, получени от мозъка, в PFC 119 , както и хроничният стрес 120 , но все още не е известно дали това допринася за наблюдаваните дендритни промени.
Хроничният стрес по време на развитието на мозъка или в детството може да има особено голям ефект върху структурата и функцията на PFC в зряла възраст. Дендритни промени могат да възникнат вътреутробно, когато майката е изложена на стрес 121 . Хроничният, неконтролируем стрес в детска възраст също може да има трайни ефекти върху PFC за възрастни. Например, малките плъхове, изложени на екстензивна майчина лишения, имат трайна дендритна ретракция в PFC и повишено поведение, подобно на тревожност 122 . За разлика от това, излагането на лек стрес, например когато непълнолетна маймуна научи, че майката ще се завърне след кратък период на раздяла, изглежда насърчава устойчивите реакции на стрес в зряла възраст 123 и намалява експресията на глюкокортикоидни рецептори в дорзолатералния PFC124 . Доказано е, че подобни ефекти се наблюдават при плъхове, при които много лекият стрес в началото на живота насърчава устойчивостта, докато по-тежките стресови фактори изострят стресовата реакция по-късно в живота 125 . Промените в утробата и по време на детството вероятно допринасят за податливостта към формални психични заболявания, които се появяват през юношеството и зрелостта 126 .
Генетични и екологични фактори
Както генетичните, така и факторите на околната среда могат да нарушат регулирането на сигналните пътища за стрес в PFC. Те могат да отслабят способностите на PFC и в някои случаи да доведат до симптоми на психично заболяване.
Генетични обиди
Някои генетични вариации в гени, които кодират ензими, които катаболизират катехоламините, които водят до относително фини ефекти върху функцията на PFC. Например, заместването на метионин в COMT увеличава стресовата реакция при мишки 75 и прави хората по-податливи на когнитивно увреждане след стрес и психостимулиращи лекарства 74 . По същия начин, генетична вариация в промоторния регион за гена на моноаминооксидаза А ( MAOA ) увеличава асоциалното поведение при момчета, изложени на стреса от насилие над деца 127 .
Промените в гените, които кодират молекули, които служат като вътреклетъчни спирачки на сигналните пътища за стрес, са свързани със сериозни психични заболявания (Фиг. 3). Например, нарушена при шизофрения 1 ( DISC1 ) обикновено осигурява отрицателна обратна връзка за cAMP сигнализиране, но мутация в DISC1 намалява активността на фосфодиестеразата PDE4B в присъствието на високи концентрации на cAMP 128 . транслокациите със загуба на функция в DISC1 са свързани с високи нива на психични заболявания 129 и с намалено PFC сиво вещество и нарушена работна памет при пациенти с шизофрения 130 . DISC1 е важен за развитието на PFC 131 , но също така е вероятно загубата на функция на DISC1 да доведе до повишено cAMP-HCN сигнализиране в бодли 132и по този начин до прекъсване на мрежата на PFC. Друга важна молекулярна спирачка на сигналните пътища за стрес е регулаторът на сигнализирането на G-протеин 4 ( RGS4 ), който инхибира G q сигнализирането. RGS4 обикновено е концентриран в дендритния ствол, където се генерират вълни Ca 2+, които изключват изгарянето на PFC клетки 133 . Нивата на RGS4 са значително намалени в PFC на пациенти с шизофрения 134 , 135 и има данни за генетични промени в RGS4 в някои семейства с шизофрения или биполярно разстройство 136 , 137. загубата на RGS4 функция би дезинхибирала фосфатидилинозитол-PKC сигнализиране, което би довело до намалено PFC стрелба на невроните и чувствителност към загуба на гръбначния стълб. И накрая, DAG киназа-η ( DGKH ) е идентифицирана като ген, който е най-силно свързан с биполярно разстройство 138 . DGK е ензимът, който хидролизира DAG и спира активирането на PKC. Важно е, че лекарствата за тези заболявания индиректно инхибират PKC сигналния път (например, литийът потиска сигнализацията на фосфатидилинозитол), а атипичните антипсихотични лекарства блокират α 1 -адренергични и серотонин (5-хидрокситриптамин, 5-НТ) тип 2 (5-HT 2 ) рецептори, които са свързани с G q сигнализиране 139 , 140. Има скорошни доказателства, че литийът може да възстанови PFC активността и нивата на сивото вещество при пациенти с биполярно разстройство 141 - 143 . По този начин лекарствата, които коригират последствията от генетични обиди, могат да възстановят нормалната структура и функция.
Екологични обиди
Стресорите понякога са под формата на екологични обиди, които застрашават нашата безопасност. излагането на травматични стресови фактори може да преувеличи последващите стресови реакции и да доведе до PTSD. PTSD се свързва с намалена медиална активност на PFC и обем на сивото вещество 144 и с повишена активност на норадреналин 145 . Образните изследвания показват, че изпитването на свързани с травма разсейващи стимули нарушава мрежите на работещата памет при пациенти с PTSD 146 . Подобни ефекти са наблюдавани при пациенти с депресия 147 . Интересното е, че жените след пубертета преди менопаузата са изложени на много по-висок риск от ПТСР и депресия 148и непокътнатите женски плъхове (т.е. плъхове с нормални нива на естроген) са по-чувствителни към стрес-индуцирана дисфункция на PFC, отколкото мъжките плъхове или яйчниците от яйчници 34 , 149 . Това предполага, че естрогенът може да усили реакцията на PFC към екологични обиди.
Различни токсини могат да увредят функцията на PFC, като преувеличават активността на сигналните пътища за стрес. Например, злоупотреби с наркотици като кокаин произвеждат прекомерна допаминова сигнализация в PFC, което отслабва PFC функцията 150 , включително способността да се противопоставят на приемането на наркотици. Това предизвиква порочен кръг, тъй като последващият прием на наркотици води до все по-слаба регулация на поведението на PFC. Приемането на наркотици е особено разпространено по време на стрес, когато функцията на PFC е допълнително отслабена от сигналните пътища за стрес 22 .
Друг важен токсин за околната среда е оловото. оловото е двувалентен катион, който имитира Ca 2+ и мощно активира PKC сигнализиране 151 . Прилагането на ниски нива на олово при плъхове води до дефицит в регулирането на вниманието на PFC, който имитира симптомите на разстройство с хипер активност с дефицит на вниманието (ADHD) 152 . При хората отравянето с олово е свързано с намаляване на PFC сивото вещество 153 и с различни поведенчески промени, които произтичат от дисфункция на PFC, включително подобни на ADHD проблеми и контрол на импулсите и дори социопатично, безотговорно и престъпно поведение 154 , 155 . отравянето с олово силно корелира с нивата на престъпност и извънбрачната бременност в съвременна Северна Америка154 , 155 . Интересното е, че се предполага, че отравянето с олово е било причина за падането на Римската империя, тъй като заможният клас е ял храни, консервирани в оловен сироп, и е ял от оловни прибори 156 . Лошото вземане на решения и импулсивното поведение на богатата класа - в съответствие с дисфункцията на PFC - доведоха до краха на велика империя 156 . По този начин фактори, които нарушават регулирането на сигналните пътища за стрес и отслабват структурата и функцията на PFC, могат да подкопаят цивилизованото поведение на цели култури.
Заключения и бъдещи указания
Огромният напредък беше постигнат в разбирането на неврохимичните влияния върху префронталната функция от първото откритие на Брозоски и колегите му преди 30 години 157 , което показа, че допаминът е от съществено значение за дорзолатералните способности на префронталната работна памет. Все още трябва да се свърши огромно количество работа, включително проучвания на допълнителни невромодулатори, различни префронтални области и други когнитивни операции на PFC. Например, тече важна работа за разкриване на сложните и мощни ефекти върху префронталните функции на серотонина, друг модулатор, който се освобождава в PFC по време на излагане на неконтролируем стрес 158 и който е ключов за орбиталната PFC функция 159. Серотонинът е особено важен за етиологията и лечението на депресията, а генетичните промени в серотониновите рецептори или серотониновия транспортер могат да повлияят на лимбичните връзки на PFC 160 . Въпреки това, малко се знае за това как серотонинът променя стрелбата на PFC клетки при животни, изпълняващи задачи, които включват PFC. едно изследване показва, че високи нива на 5-НТ 2А рецепторна стимулация може да потисне дорзолатералното PFC невронална удар маймуни изпълняващи задача работната памет 161 . Тези резултати са подобни на тези, наблюдавани с високи нива на стимулиране на α 1 адренергичен рецептор, които, като 5-НТ 2Арецептор, се свързва с фосфатидилинозитол-PKC сигнализиране. Неотдавнашно проучване при плъхове предполага, че серотониновата стимулация на 5-НТ 2С рецепторите влошава обръщането на възнаграждението 162 , функция, която е силно зависима от орбиталния PFC. Би било очарователно да се изследва дали 5-НТ 2С рецепторна стимулация ще потисне изстрелването на орбитални PFC неврони при маймуни, изпълняващи задача за обръщане на наградата, и дали тези ефекти могат да бъдат блокирани от агенти, които инхибират сигнализирането на фосфатидилинозитол-ПКК.
Вредното въздействие на стреса върху PFC мрежите е особено проблематично в „информационната ера“, когато опосредстваните от PFC когнитивни способности са все по-необходими за успех. разбирането на молекулярните механизми, които променят структурата на PFC и нарушават функцията на PFC по време на излагане на стрес, ще помогне да се разкрие как генетичните и екологичните обиди могат да увеличат чувствителността на индивида към дефицити на PFC. Идентифицирането на молекулярните механизми, които променят функцията на PFC, ще осигури основата за нова ера в психиатрията, когато ще разберем как генетичните промени влияят върху вътреклетъчната сигнализация, невронното развитие и неврофизиологията във веригите, които са в основата на невропсихиатричните симптоми.
Благодарности
Голяма част от изследванията, цитирани в този преглед, са подкрепени от MERIT Award AG06036 и от P50MH068789, PO1AG030004 и RL1AA017536 като част от U54RR024350, както и от Института по неврология на Кавли в Йейл и Национален алианс за изследване на шизофрения и депресия Награда за AFTA
Терминологичен речник
Комплект за внимание Предразположение към едно измерение на даден стимул, като същевременно инхибира други измерения - например, внимание към цвета, а не към формата. Trier тест за социален стрес Тест, при който субектите трябва да изнесат реч и да извършват изчисления пред група от хора, които не познават. Проби от кръв или слюнка и измервания на кръвното налягане могат да се вземат преди, по време и след теста, за да се определи физическата реакция на стресора. Научена парадигма за безпомощност Парадигма, разработена преди повече от 30 години, при която плъховете са били изложени на неизбежен шок и се предполага, че са научили, че са безпомощни да реагират. Изследванията развенчаха тази интерпретация и вместо това установиха, че неконтролируемият стрес може да причини когнитивни дефицити. Задача Y-лабиринт Задача, при която плъховете трябва да се научат да избягат от Y-образния лабиринт, като вземат правилното решение. Установено е, че излагането на неконтролируем стрес и наличието на неподходящи реплики в лабиринта влошава работата. Задача за пространствено забавено редуване Тест за пространствена работна памет за примати или гризачи, при който субектът трябва да даде алтернативни отговори при последователни опити, с период на закъснение между интервалите. Настроено постоянно изстрелване на неврони Невроналното представяне на специфичен стимул - например реплика в определено пространствено местоположение. Поради мрежовите връзки клетката може да поддържа стрелба без стимулиране от околната среда, но продължителната стрелба (в този пример) се случва само след указание за определено място в космоса; по този начин невронът е „настроен“ в тази посока. Фосфодиестераза Ензим, който хидролизира сАМР. Инхибирането на фосфодиестеразата води до натрупване на концентрации на сАМР. Бележки под линия
Изявление за конкуриращи се интереси
Авторът декларира конкуриращи се финансови интереси : вижте уеб версията за подробности.
БАЗИ ДАННИ
Entrez ген: http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?db=gene
COMT | DGKH | DISC1 | EPAC | MAOA | PDE4B | RGS4
ОЩЕ ИНФОРМАЦИЯ
Началната страница на Amy ft Arnsten: http://info.med.yale.edu/neurobio/arnsten/Index.html
Препратки
1. Fuster JM. Префронталната кора. Академична преса; 2008. [ Google Scholar ] Това е най-новото издание на класическа и красноречива книга за PFC.2. Goldman-Rakic PS. Префронталният пейзаж: последици от функционалната архитектура за разбиране на човешката ментация и централната изпълнителна власт. Филос. Транс. R. Soc. Лонд. B Biol. Sci. 1996; 351 : 1445–1453. [ PubMed ] [ Google Scholar ]3. Thompson-Schill SL, et al. Ефекти от увреждане на фронталния лоб върху смущаващи ефекти в работната памет. Когн. Афект. Behav. Невроски. 2002; 2 : 109–120. [ PubMed ] [ Google Scholar ]4. Aron AR, Robbins TW, Poldrack RA. Инхибиране и дясната долна челна кора. Тенденции Когн. Sci. 2004; 8 : 170–177. [ PubMed ] [ Google Scholar ]5. Buschman TJ, Miller EK. Контрол на вниманието отгоре надолу спрямо дъното нагоре в префронталната и задната теменна кора. Наука. 2007; 315 : 1860–1862. [ PubMed ] [ Google Scholar ]6. Gazzaley A, et al. Функционалните взаимодействия между префронталната и зрителната кора на асоциацията допринасят за модулация на визуалната обработка отгоре надолу. Цереб. Кортекс. 2007; 17 (Допълнение 1): i125 – i135. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]7. Робинс TW. От възбуда до познание: интегративното положение на префронталната кора. Prog. Brain Res. 2000; 126 : 469–483. [ PubMed ] [ Google Scholar ] Този преглед обединява важна информация за предфронталните вериги и как те са модулирани.8. Lee D, Seo H. Механизми на засилване на обучение и вземане на решения в приморската дорсолатерална префронтална кора. Ан. Ню Йорк акад. Sci. 2007; 1104 : 108–122. [ PubMed ] [ Google Scholar ]9. Modirrousta M, Fellows LK. Дорзалната медиална префронтална кора играе необходима роля за бързото прогнозиране на грешки при хората. J. Neurosci. 2008; 28 : 14000–14005. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]10. Broadbent D. Решение и стрес. Лондон: Академик; 1971. [ Google Scholar ] Това е класическа книга за ефектите от стреса върху когнитивните функции.11. Хокей GRJ. Ефект на силен шум върху внимателността на селективността. QJ опит. Психол. 1970; 22 : 28–36. [ Google Scholar ]12. Хартли Л.Р., Адамс Р.Г. Ефект на шума върху теста на Stroop. J. опит. Психол. 1974; 102 : 62–66. [ PubMed ] [ Google Scholar ]13. Arnsten AFT. Биологията на чувството за раздразнение. Наука. 1998; 280 : 1711–1712. [ PubMed ] [ Google Scholar ]14. Elliott AE, Packard MG. Интраамигдална анксиогенна инфузия на лекарства преди извличане пристрастява плъховете към използването на навик памет. Невробиол. Уча. Мем. 2008; 90 : 616–623. [ PubMed ] [ Google Scholar ]15. Стъклен DC, Reim B, Singer JE. Поведенчески последици от адаптацията към контролируем и неконтролируем шум. J. опит. Социален психолог. 1971; 7 : 244–257. [ Google Scholar ]16. Миньор TR, Джаксън RL, Maier SF. Ефекти от релевантни на задачите реплики и забавяне на усилването върху обучението за избягване след неизбежен шок: доказателства за дефицит в селективното внимание. J. опит. Психол. Anim. Behav. Процес. 1984; 10 : 543–556. [ PubMed ] [ Google Scholar ]17. Qin S, Hermans EJ, van Marle HJF, Lou J, Fernandez G. Остър психологически стрес намалява свързаната с работната памет активност в дорзолатералната префронтална кора. Biol. Психиатрия. (в пресата). [ PubMed ] [ Google Scholar ]18. Dolcos F, McCarthy G. Мозъчни системи, медииращи когнитивната намеса чрез емоционално разсейване. J. Neurosci. 2006; 26 : 2072–2079. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]19. Александър JK, Hillier A, Smith RM, Tivarus ME, Beversdorf DQ. Бета-адренергична модулация на когнитивната гъвкавост по време на стрес. J. Cogn. Невроски. 2007; 19 : 468–478. [ PubMed ] [ Google Scholar ]20. Luethi M, Meier B, Sandi C. Стресови ефекти върху работната памет, явна памет и имплицитна памет за неутрални и емоционални стимули при здрави мъже. Отпред. Behav. Невроски. 2009 г. 15 януари; (doi: 10.3389 / neuro.08.005.2008). [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]21. Sinha R, Lacadie CM, Skudlarski P, Wexler BE. Невронни вериги, лежащи в основата на емоционален дистрес при хората. Ан. Ню Йорк акад. Sci. 2004; 1032 : 254–257. [ PubMed ] [ Google Scholar ]22. Li CS, Sinha R. Инхибиторен контрол и регулиране на емоционалния стрес: невроизобразителни доказателства за фронтално-лимбична дисфункция при психостимулантна зависимост. Невроски. Биобехав. Rev. 2008; 32 : 581–597. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ] Тази статия свързва префронталната дисфункция по време на стрес със злоупотребата с вещества.23. Liston C, McEwen BS, Casey BJ. Психосоциалният стрес обратимо нарушава префронталната обработка и контрола върху вниманието. Proc. Natl Акад. Sci. САЩ. 2009; 106 : 912–917. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ] Тази статия включва данни за това как хроничният стрес отслабва префронталната свързаност както при хора, така и при плъхове, изложени на хроничен стрес.24. Mazure CM, редактор. Стресът причинява ли психиатрични заболявания? Вашингтон: Американска психиатрична преса; 1995. [ Google Scholar ] Тази книга дава много примери за това как стресът влошава психичните заболявания.25. Mazure CM, Maciejewski PK. Модел на риск от голяма депресия: ефектите от житейския стрес и когнитивния стил варират в зависимост от възрастта. Депресия. Безпокойство. 2003; 17 : 26–33. [ PubMed ] [ Google Scholar ]26. Southwick S, Rasmusson A, Barron X, Arnsten AFT. В: Невропсихология на ПТСР: Биологични, когнитивни и клинични перспективи. Vasterling JJ, Brewin CR, редактори. Ню Йорк: Guilford Publications; 2005. с. 27–58. [ Google Scholar ]27. Breier A, Wolkowitz O, Pickar D. In: Шизофрения Изследвания. Tamminga C, Schult S, редактори. Кн. 1. Ню Йорк: Raven; 1991. [ Google Scholar ]28. Dohrenwend BP, Shrout PE, Link BG, Skodol AE, Stueve A. In: Стресът причинява ли психиатрични заболявания? Mazure CM, редактор. Вашингтон: Американска психиатрична преса; 1995. с. 43–65. [ Google Scholar ]29. Hammen C, Gitlin M. Стресова реактивност при биполярни пациенти и връзката му с анамнеза за разстройство. Am. J. Психиатрия. 1997; 154 : 856–857. [ PubMed ] [ Google Scholar ]30. Amat J, Paul E, Zarza C, Watkins LR, Maier SF. Предишен опит с поведенчески контрол над стреса блокира поведенческите и дорзалните ядра на рафето, активиращи ефектите на по-късно неконтролируем стрес: роля на вентралната медиална префронтална кора. J. Neurosci. 2006; 26 : 13264–13272. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ] Това проучване показа, че PFC е този, който установява (правилно или погрешно) дали контролираме стресора. PFC потиска реакцията на мозъчния ствол на стрес, дори когато възприеманият контрол е само илюзия.31. Diorio D, Viau V, Meaney MJ. Ролята на медиалната префронтална кора (cingulate gyrus) в регулирането на хипоталамо-хипофизарно-надбъбречните реакции на стрес. J. Neurosci. 1993; 13 : 3839–3847. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]32. Murphy BL, Arnsten AFT, Goldman-Rakic PS, Roth RH. Повишеният оборот на допамина в префронталната кора нарушава работата на пространствената работна памет при плъхове и маймуни. Proc. Natl Акад. Sci. САЩ. 1996; 93 : 1325–1329. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]33. Arnsten AFT, Goldman-Rakic PS. Шумовият стрес влошава префронталната кортикална когнитивна функция при маймуните: доказателства за хипердопаминергичен механизъм. Арх. Генерал Психиатрия. 1998; 55 : 362–369. [ PubMed ] [ Google Scholar ]34. Шански Р.М., Рубинов К, Бренан А, Арнстен АФ. Ефектите от секса и хормоналния статус върху увреждането на работната памет, предизвикано от стрес. Behav. Мозъчна функция. 2006; 2 : 8. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]35. McEwen BS. Защита и увреждане от остър и хроничен стрес: алостаза и алостатично претоварване и значение за патофизиологията на психиатричните разстройства. Ан. Ню Йорк акад. Sci. 2004; 1032 : 1–7. [ PubMed ] [ Google Scholar ]36. Cahill L, McGaugh JL. Модулация на паметта. Curr. Становище. Невробиол. 1996; 6 : 237–242. [ PubMed ] [ Google Scholar ]37. Quirk GJ, Mueller D. Невронни механизми на учене и извличане на изчезване. Невропсихофармакология. 2008; 33 : 56–72. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]38. Ким Джей Джей, Йон К.С. Стрес: метапластични ефекти в хипокампуса. Тенденции Neurosci. 1998; 21 : 505–509. [ PubMed ] [ Google Scholar ]39. Packard MG, Teather LA. Модулация на амигдала на множество системи с памет: хипокампус и опашни путамени. Невробиол. Уча. Мем. 1998; 69 : 163–203. [ PubMed ] [ Google Scholar ]40. Голдман-Ракич П.С. Клетъчна основа на работната памет. Неврон. 1995; 14 : 477–485. [ PubMed ] [ Google Scholar ] Тази статия описва микросхемите, които са в основата на пространствената работна памет, обобщавайки както тяхната анатомия, така и тяхната физиология.41. Funahashi S, Bruce CJ, Goldman-Rakic PS. Мнемонично кодиране на зрителното пространство в дорзолатералната префронтална кора на маймуната. J. Неврофизиол. 1989; 61 : 331–349. [ PubMed ] [ Google Scholar ]42. Aston-Jones G, Rajkowski J, Cohen J. Роля на locus coeruleus в вниманието и поведенческата гъвкавост. Biol. Психиатрия. 1999; 46 : 1309–1320. [ PubMed ] [ Google Scholar ]43. Шулц В. Фазичният сигнал за възнаграждение на допаминовите неврони на приматите. Adv. Pharmacol. 1998; 42 : 686–690. [ PubMed ] [ Google Scholar ]44. Matsumoto M, Hikosaka O. Възбуждащи и инхибиторни реакции на допаминовите неврони на средния мозък към сигнали, предсказващи нежелани стимули. Soc. Невроски. Абстракция 2008; 691,24 [ Google Scholar ]45. Roth RH, Tam SY, Ida Y, Yang JX, Deutch AY. Стресът и мезокортиколимбичните допаминови системи. Ан. Ню Йорк акад. Sci. 1988; 537 : 138–147. [ PubMed ] [ Google Scholar ]46. Finlay JM, Zigmond MJ, Abercrombie ED. Повишено освобождаване на допамин и норепинефрин в медиалната префронтална кора, предизвикано от остър и хроничен стрес: ефекти на диазепама. Неврология. 1995; 64 : 619–628. [ PubMed ] [ Google Scholar ]47. Deutch AY, Roth RH. Детерминантите на предизвиканото от стрес активиране на префронталната кортикална допаминова система. Prog. Brain Res. 1990; 85 : 367–403. [ PubMed ] [ Google Scholar ] Тази статия обобщава многобройните биохимични изследвания на освобождаването на допамин в PFC по време на излагане на стрес.48. Lewis DA, Cambell MJ, Foote SL, Goldstein M, Morrison JH. Разпределението на тирозин хидроксилаза-имунореактивни влакна в неокортекса на приматите е широко разпространено, но регионално специфично. J. Neurosci. 1987; 282 : 317–330. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]49. McCormick DA, Pape HC, Williamson A. Действия на норадреналина в мозъчната кора и таламуса: последици за функцията на централната норадренергична система. Prog. Brain Res. 1991; 88 : 293–305. [ PubMed ] [ Google Scholar ]50. Seamans JK, Durstewitz D, Christie BR, Stevens CF, Sejnowski TJ. Допаминова D1 / D5 рецепторна модулация на възбуждащи синаптични входове към V неврони на префронталната кора на слой V. Proc. Natl Акад. Sci. САЩ. 2001; 98 : 301–306. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]51. Арнстен АФТ. През огледалото: диференциална норадренергична модулация на префронталната функция на кората. Neural Plast. 2000; 7 : 133–146. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]52. Arnsten AFT, Goldman-Rakic PS. Алфа-2 адренергични механизми в префронталната кора, свързани с когнитивен спад при възрастни нечовешки примати. Наука. 1985; 230 : 1273–1276. [ PubMed ] [ Google Scholar ]53. Li BM, Mei ZT. Дефицит на забавен отговор, предизвикан от локално инжектиране на алфа-2 адренергичния антагонист йохимбин в дорзолатералната префронтална кора при млади възрастни маймуни. Behav. Neural Biol. 1994; 62 : 134–139. [ PubMed ] [ Google Scholar ]54. Birnbaum SG, Gobeske KT, Auerbach J, Taylor JR, Arnsten AFT. Роля на норепинефрин при когнитивни дефицити, предизвикани от стрес: медиация на α-1-адренорецептора в префронталната кора. Biol. Психиатрия. 1999; 46 : 1266–1274. [ PubMed ] [ Google Scholar ]55. Ramos B, et al. Бета-1 адренергичният антагонист, бетаксолол, подобрява работата на работната памет при плъхове и маймуни. Biol. Психиатрия. 2005; 58 : 894–900. [ PubMed ] [ Google Scholar ]56. Cai JX, Ma Y, Xu L, Hu X. Резерпината влошава ефективността на пространствената работна памет при маймуните: обръщане от алфа-2 адренергичния агонист клонидин. Brain Res. 1993; 614 : 191–196. [ PubMed ] [ Google Scholar ]57. Мао ZM, Arnsten AFT, Li BM. Локалната инфузия на алфа-1 адренергичен агонист в префронталната кора нарушава работата на пространствената работна памет при маймуните. Biol. Психиатрия. 1999; 46 : 1259–1265. [ PubMed ] [ Google Scholar ]58. Ramos B, Stark D, Verduzco L, van Dyck CH, Arnsten AFT. Алфа-2А-адренорецепторната стимулация подобрява префронталната кортикална регулация на поведението чрез инхибиране на cAMP сигнализирането при застаряващи животни. Уча. Мем. 2006; 13 : 770–776. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]59. Li BM, Mao ZM, Wang M, Mei ZT. Алфа-2 адренергична модулация на префронталната кортикална невронална активност, свързана с пространствената работна памет при маймуни. Невропсихофармакология. 1999; 21 : 601–610. [ PubMed ] [ Google Scholar ]60. Wang M, et al. Стимулирането на α2A-адренорецепторите укрепва мрежите на работещата памет, като инхибира сигнализирането на cAMP-HCN канал в префронталната кора. Клетка. 2007; 129 : 397–410. [ PubMed ] [ Google Scholar ]61. Birnbaum SB, et al. Свръхактивността на протеин киназа С нарушава префронталната кортикална регулация на работната памет. Наука. 2004; 306 : 882–884. [ PubMed ] [ Google Scholar ]62. Arnsten AFT, Mathew R, Ubriani R, Taylor JR, Li BM. Стимулирането на α-1 норадренергичен рецептор нарушава когнитивната функция на префронталната кора. Biol. Психиатрия. 1999; 45 : 26–31. [ PubMed ] [ Google Scholar ]63. Тейлър Ф, Раскинд М.А. А1-адренергичният антагонист празозин подобрява съня и кошмарите при цивилно травматично посттравматично стресово разстройство. J. Clin. Психофармакол. 2002; 22 : 82–85. [ PubMed ] [ Google Scholar ]64. Raskind MA, et al. Празозин намалява кошмарите и други симптоми на ПТСР при бойни ветерани: плацебо-контролирано проучване. Am. J. Психиатрия. 2003; 160 : 371–373. [ PubMed ] [ Google Scholar ]65. Arnsten AFT, Goldman-Rakic PS. Стресът нарушава когнитивната функция на префронталната кора при маймуните: роля на допамина. Soc. Невроски. Абстракция 1990; 16 : 164. [ Google Scholar ]66. Sawaguchi T, Goldman-Rakic PS. D1 допаминови рецептори в префронталната кора: участие в работната памет. Наука. 1991; 251 : 947–950. [ PubMed ] [ Google Scholar ]67. Zahrt J, Taylor JR, Mathew RG, Arnsten AFT. Супранормалната стимулация на допаминови D1 рецептори в префронталната кора на гризачите нарушава работата на пространствената работна памет. J. Neurosci. 1997; 17 : 8528–8535. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]68. Vijayraghavan S, et al. Действия на инвертирания U допамин D1 рецептор върху префронтални неврони, ангажирани в работната памет. Nature Neurosci. 2007; 10 : 376–384. [ PubMed ] [ Google Scholar ]69. Друзин М.Й., Курзина Н.П., Малинина Е.П., Козлов А.П. Ефектите от локалното приложение на D2 селективни допаминергични лекарства в медиалната префронтална кора на плъхове при задача за забавен пространствен избор. Behav. Brain Res. 2000; 109 : 99–111. [ PubMed ] [ Google Scholar ]70. Gibbs SE, D'Esposito M. Функционално ЯМР изследване на ефектите на бромокриптин, агонист на допаминовия рецептор, върху компонентни процеси на работната памет. Психофармакология. 2005; 180 : 644–653. [ PubMed ] [ Google Scholar ]71. Wang M, Vijayraghavan S, Goldman-Rakic PS. Селективни действия на D2 рецептора върху функционалната схема на работната памет. Наука. 2004; 303 : 853–856. [ PubMed ] [ Google Scholar ]72. Kimberg DY, D'Esposito M, Farah MJ. Ефектите на бромокриптин върху хора зависят от капацитета на работната памет. Неврорепортаж. 1997; 8 : 3581–3585. [ PubMed ] [ Google Scholar ]73. Egan MF, et al. Ефект на COMT Val 108/158 Met генотип върху функцията на фронталния лоб и риск от шизофрения. Proc. Natl Акад. Sci. САЩ. 2001; 98 : 6917–6922. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]74. Mattay VS, et al. Катехол O-метилтрансфераза val158-met генотип и индивидуални вариации в мозъчния отговор на амфетамин. Proc. Natl Акад. Sci. САЩ. 2003; 100 : 6186–6191. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]75. Papaleo F, et al. Генетична дисекция на ролята на катехол-О-метилтрансферазата в когнитивността и реактивността на стреса при мишки. J. Neurosci. 2008; 28 : 8709–8723. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]76. Roozendaal B, McReynolds JR, McGaugh JL. Базолатералната амигдала взаимодейства с медиалната префронтална кора при регулиране на глюкокортикоидните ефекти върху увреждането на работната памет. J. Neurosci. 2004; 24 : 1385–1392. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]77. Lupien SJ, Gillin CJ, Hauger RL. Работната памет е по-чувствителна от декларативната памет към острите ефекти на кортикостероидите: проучване на доза-отговор при хора. Behav. Невроски. 1999; 113 : 420–430. [ PubMed ] [ Google Scholar ]78. Grundemann D, Schechinger B, Rappold GA, Schomig E. Молекулярна идентификация на чувствителния към кортизон екстраневронален катехоламинов транспортер. Nature Neurosci. 1998; 1 : 349–351. [ PubMed ] [ Google Scholar ]79. Ферибот B, Roozendaal B, McGaugh JL. Базолатералните амигдални норадренергични влияния върху паметта се медиират от взаимодействие между β- и α-1-адренорецепторите. J. Neurosci. 1999; 19 : 5119–5123. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]80. Roozendaal B, Quirarte GL, McGaugh JL. Глюкокортикоидите взаимодействат с базолатералната амигдала β-адренорецептор – cAMP / cAMP / PKA система при въздействие върху консолидирането на паметта. Евро. J. Neurosci. 2002; 15 : 553–560. [ PubMed ] [ Google Scholar ]81. Goldstein LE, Rasmusson AM, Bunney SB, Roth RH. Роля на амигдалата в координацията на поведенчески, невроендокринни и префронтални кортикални моноаминови реакции на психологически стрес при плъхове. J. Neurosci. 1996; 16 : 4787–4798. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]82. Hopkins WF, Johnston D. Норадренергично усилване на дългосрочното потенциране при мъхообразни влакнести синапси в хипокампуса. J. Neurophys. 1988; 59 : 667–687. [ PubMed ] [ Google Scholar ]83. Hu H, et al. Емоцията подобрява ученето чрез регулиране на норепинефрин на трафика на AMPA-рецептори. Клетка. 2007; 131 : 160–173. [ PubMed ] [ Google Scholar ]84. Foote SL, Freedman FE, Oliver AP. Ефекти на предполагаемите невротрансмитери върху невроналната активност в слухова кора на маймуни. Brain Res. 1975; 86 : 229–242. [ PubMed ] [ Google Scholar ]85. Waterhouse BD, Moises HC, DJ Woodward. Норадренергична модулация на соматосензорни кортикални невронални отговори към йонофоретично приложени предполагаеми предаватели. Опит Неврол. 1980; 69 : 30–49. [ PubMed ] [ Google Scholar ]86. Waterhouse BD, Moises HC, DJ Woodward. Алфа-рецептор-медиирано улесняване на соматосензорни кортикални невронални отговори на възбуждащи синаптични входове и йонофоретично прилаган ацетилхолин. Неврофармакология. 1981; 20 : 907–920. [ PubMed ] [ Google Scholar ]87. Wickens JR, Horvitz JC, Costa RM, Killcross S. Допаминергични механизми в действия и навици. J. Neurosci. 2007; 27 : 8181–8183. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]88. Runyan JD, Moore AN, Dash PK. Роля за активността на префронталната чувствителна на калций протеинова фосфатаза и киназа в работната памет. Уча. Мем. 2005; 12 : 103–110. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]89. Hagenston AM, Fitzpatrick JS, Yeckel MF. mGluR-медиираните калциеви вълни, които нахлуват в сомата, регулират стрелбата в медиален слой V префронтални кортикални пирамидални неврони. Цереб. Кортекс. 2008; 18 : 407–423. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]90. Runyan JD, Dash PK. Различни префронтални молекулярни механизми за съхранение на информация с продължителност секунди спрямо минути. Уча. Мем. 2005; 12 : 232–238. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ] Това важно проучване демонстрира, че отделните процеси на паметта могат да бъдат модулирани по много различни начини, дори в един и същ мозъчен регион.91. Bos JL. Epac протеини: многоцелеви сАМР цели. Тенденции Biochem. Sci. 2006; 31 : 680–686. [ PubMed ] [ Google Scholar ]92. Wahl-Schott C, Biel M. HCN канали: структура, клетъчна регулация и физиологична функция. Клетка. Мол. Life Sci. 2009; 66 : 470–494. [ PubMed ] [ Google Scholar ]93. Chen S, Wang J, Siegelbaum SA. Свойства на активирания от хиперполяризация ток на пейсмейкъра, дефиниран чрез съвместно сглобяване на субединици HCN1 и HCN2 и базална модулация от цикличен нуклеотид. J. Gen. Physiol. 2001; 117 : 491–504. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]94. Джордж MS, Abbott LF, Siegelbaum SA. Активираните от хиперполяризация HCN канали инхибират подпраговите EPSP чрез зависими от напрежението взаимодействия с М + тип К + канали. Nature Neurosci. (в пресата). [ Безплатна статия за PMC ] [ PubMed ] [ Google Scholar ] Това важно ново проучване използва както изчислително моделиране, така и физиология, за да обясни как HCN каналите могат да шунтират входящите синаптични входове.95. Delmas P, Brown DA. Пътища, модулиращи невронни KCNG / M (Kv7) калиеви канали. Nature Rev. Neurosci. 2005; 6 : 850–862. [ PubMed ] [ Google Scholar ]96. Taylor JR, Birnbaum SG, Ubriani R, Arnsten AFT. Активирането на протеин киназа А в префронталната кора нарушава работата на работната памет. J. Neurosci. 1999; 19 : RC23. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]97. Доктор Сулсби, RJ Wojcikiewicz. 1,4,5-трисфосфатният рецептор на инозитол тип III се фосфорилира от сАМР-зависима протеин киназа на три места. Biochem. J. 2005; 392 : 493–497. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]98. Ferguson GD, Storm DR. Защо калциево стимулирани аденилил циклази? Физиология. 2004; 19 : 271–276. [ PubMed ] [ Google Scholar ]99. Яребица LD, Swandulla D, Muller TH. Модулация на калциево активирани неспецифични катионни токове чрез циклично AMP-зависимо фосфорилиране в неврони на Helix . J. Physiol. 1990; 429 : 131–145. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]100. Soboloff J, et al. TRPC канали: интегратори на множество клетъчни сигнали. Дръжка. опит. Pharmacol. 2007; 179 : 575–591. [ PubMed ] [ Google Scholar ]101. Haj-Dahmane S, Andrade R. Йонен механизъм на бавната адепополяризация, индуцирана от активирането на мускариновия рецептор в префронталната кора на плъхове. J. Неврофизиол. 1998; 80 : 1197–1210. [ PubMed ] [ Google Scholar ]102. Tegner J, Compte A, Wang XJ. Динамичната стабилност на реверберационните невронни вериги. Biol. Киберн. 2002; 87 : 471–481. [ PubMed ] [ Google Scholar ]103. Han JZ, Lin W, Lou SJ, Qiu J, Chen YZ. Бързо, негеномно действие на глюкокортикоиди в клетки на невробластом B103 на плъх. Biochim. Biophys. Acta. 2002; 1591 : 21–27. [ PubMed ] [ Google Scholar ]104. Холмс А, Уелман CL. Индуцирана от стрес префронтална реорганизация и изпълнителна дисфункция при гризачи. Невроски. Биобехав. Rev. 2008 6 декември; (doi: 10.1016 / j. neubiorev.2008.11.005). [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]105. Radley JJ, et al. Повтарящият се стрес предизвиква дендритна загуба на гръбначния стълб в медиалната префронтална кора на плъховете. Цереб. Кортекс. 2006; 16 : 313–320. [ PubMed ] [ Google Scholar ]106. Michelsen KA, et al. Пренаталният стрес и последващото излагане на хроничен лек стрес оказват влияние върху дендритната плътност на гръбначния стълб и морфологията в медиалната префронтална кора на плъховете. BMC Neurosci. 2007; 8 : 107. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]107. Liston C, et al. Индуцираните от стреса промени в префронталната кортикална дендритна морфология предсказват селективни увреждания при перцептивното преместване на вниманието. J. Neurosci. 2006; 26 : 7870–7874. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]108. Cerqueira JJ, Mailliet F, Almeida OF, Jay TM, Sousa N. Префронталната кора като ключова цел на дезадаптивния отговор на стреса. J. Neurosci. 2007; 27 : 2781–2787. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]109. Radley JJ, et al. Обратимост на апикалната дендритна ретракция в медиалната префронтална кора на плъх след повтарящ се стрес. Опит Неврол. 2005; 196 : 199–203. [ PubMed ] [ Google Scholar ]110. Браун SM, Henning S, Wellman CL. Лекият, краткосрочен стрес променя дендритната морфология в медиалната префронтална кора на плъховете. Цереб. Кортекс. 2005; 15 : 1714–1722. [ PubMed ] [ Google Scholar ]111. Izquierdo A, Wellman CL, Holmes A. Кратък неконтролируем стрес причинява дендритна ретракция в инфралимбичната кора и устойчивост на изчезване на страх при мишки. J. Neurosci. 2006; 26 : 5733–5738. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]112. Vyas A, Mitra R, Shankaranarayana Rao BS, Chattarji S. Хроничният стрес предизвиква контрастни модели на дендритно ремоделиране в хипокампални и амигдалоидни неврони. J. Neurosci. 2002; 22 : 6810–6818. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]113. Shansky RM, Hamo C, Hof PR, McEwen BS, Morrison JH. Индуцираното от стрес дендритно ремоделиране в префронталната кора е специфично за веригата. Цереб. Кортекс. 2009 г. 4 февруари; (doi: 10.1093 / cercor / bhp003). [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ] Това проучване показва, че не всички префронтални неврони реагират по подобен начин на стрес и че има отделни промени въз основа на включените вериги.114. Miner LH, et al. Хроничният стрес увеличава плазменото разпределение на норепинефриновия транспортер и коекспресията на тирозин хидроксилаза в норепинефриновите аксони в префронталната кора. J. Neurosci. 2006; 26 : 1571–1578. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]115. Mizoguchi K, et al. Хроничният стрес предизвиква увреждане на пространствената работна памет поради префронтална допаминергична дисфункция. J. Neurosci. 2000; 20 : 1568–1575. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]116. Wellman CL. Дендритна реорганизация в пирамидални неврони в медиалната префронтална кора след хронично приложение на кортикостерон. J. Neurobiol. 2001; 49 : 245–253. [ PubMed ] [ Google Scholar ]117. Liu RJ, Aghajanian GK. Стресът притъпява предизвиканите от серотонин и хипокретин EPSCs в префронталната кора: роля на кортикостерон-медиирана апикална дендритна атрофия. Proc. Natl Акад. Sci. САЩ. 2008; 105 : 359–364. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]118. Zhu MY, Wang WP, Huang J, Regunathan S. Хроничното лечение с глюкокортикоиди променя морфологията на хипокампалната и префронталната кортикална морфология на плъхове паралелно с нивата на ендогенни агматин и аргинин декарбоксилаза. J. Neurochem. 2007; 103 : 1811–1820. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]119. Gourley SL, Kedves AT, Olausson P, Taylor JR. Историята на експозиция на кортикостерон регулира изчезването на страха и кортикалните NR2B, GluR2 / 3 и BDNF. Невропсихофармакология. 2009; 34 : 707–716. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]120. Lin Y, et al. Полови различия в ефектите на остър и хроничен стрес и възстановяване след дълготраен стрес върху свързаните със стреса мозъчни области на плъхове. Цереб. Кортекс. 2008 г. 10 декември; (doi: 10.1093 / cercor / bhn225). [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]121. Murmu MS, et al. Промени в плътността на гръбначния стълб и дендритната сложност в префронталната кора при потомството на майки, изложени на стрес по време на бременност. Евро. J. Neurosci. 2006; 24 : 1477–1487. [ PubMed ] [ Google Scholar ]122. Pascual R, Замора-Леон SP. Ефекти на неонаталната майчина депривация и сложността на околната среда след отбиването върху дендритната морфология на префронталните пирамидални неврони при плъхове. Acta Neurobiol. опит. (Войни.) 2007; 67 : 471–479. [ PubMed ] [ Google Scholar ]123. Parker KJ, Buckmaster CL, Justus KR, Schatzberg AF, Lyons DM. Лекият стрес в началото на живота засилва инхибирането на префронтално зависимия отговор при маймуните. Biol. Психиатрия. 2005; 57 : 848–855. [ PubMed ] [ Google Scholar ]124. Patel PD, Katz M, Karssen AM, Lyons DM. Индуцирани от стреса промени в експресията на кортикостероидни рецептори в хипокампуса на приматите и префронталната кора. Психоневроендокринология. 2008; 33 : 360–367. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]125. Meaney MJ, et al. Ранно регулиране на околната среда на експресията на ген на глюкокортикоидни рецептори на предния мозък: последици за адренокортикалните реакции на стрес. Dev. Невроски. 1996; 18 : 49–72. [ PubMed ] [ Google Scholar ]126. Lupien SJ, McEwen BS, Gunnar MR, Heim C. Ефекти на стреса през целия живот върху мозъка, поведението и познанието. Nature Rev. Neurosci. 2009 г. 29 април; (doi: 10.1038 / nrn2639). [ PubMed ] [ Google Scholar ]127. Kim-Cohen J, et al. MAOA, малтретиране и взаимодействие с генната среда, предсказващи психичното здраве на децата: нови доказателства и мета-анализ. Мол. Психиатрия. 2006; 11 : 903–913. [ PubMed ] [ Google Scholar ]128. Millar JK, et al. DISC1 и PDE4B са взаимодействащи генетични фактори при шизофрения, които регулират cAMP сигнализирането. Наука. 2005; 310 : 1187–1191. [ PubMed ] [ Google Scholar ]129. Millar JK, et al. Нарушаване на два нови гена чрез транслокация, съ-сегрегираща се с шизофрения. Хъм. Мол. Genet. 2000; 9 : 1415–1423. [ PubMed ] [ Google Scholar ]130. Cannon TD, et al. Асоциация на хаплотипите DISC1 / TRAX с шизофрения, намалено префронтално сиво вещество и нарушена краткосрочна и дългосрочна памет. Арх. Генерал Психиатрия. 2005; 62 : 1205–1213. [ PubMed ] [ Google Scholar ]131. Ishizuka K, Paek M, Kamiya A, Sawa A. Преглед на Disrupt-In-Schizophrenia-1 (DISC1): невроразвитие, познание и психични състояния. Biol. Психиатрия. 2006; 59 : 1189–1197. [ PubMed ] [ Google Scholar ]132. Kirkpatrick B, et al. DISC1 имунореактивност на светлинно и ултраструктурно ниво в човешката неокортекс. J. Comp. Неврол. 2006; 497 : 436–450. [ PubMed ] [ Google Scholar ]133. CD Паспалас, Селемон LD, Арнстен AF. Картиране на регулатора на сигнализиране на G протеин 4 (RGS4): пресинаптични и постсинаптични субстрати за неврорегулация в префронталната кора. Цереб. Кортекс. 2009 г. 19 януари; (doi: 10.1093 / cercor / bhn235). [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]134. Mirnics K, Middleton FA, Stanwood GD, Lewis DA, Levitt P. Специфични за заболяването промени в регулатора на експресия на сигнализиране на G-протеин 4 (RGS4) при шизофрения. Мол. Психиатрия. 2001; 6 : 293–301. [ PubMed ] [ Google Scholar ]135. Erdely HA, Tamminga CA, Roberts RC, Vogel MW. Регионални промени в протеина RGS4 при шизофрения. Синапс. 2006; 59 : 472–479. [ PubMed ] [ Google Scholar ]136. Chowdari KV, et al. Анализ на асоциирането и връзката на полиморфизмите RGS4 при шизофрения. Хъм. Мол. Genet. 2002; 11 : 1373–1380. [ PubMed ] [ Google Scholar ]137. Morris DW, et al. Потвърждаване на RGS4 като ген за чувствителност към шизофрения. Am. J. Med. Genet. B Невропсихиатър. Genet. 2004; 125 : 150–153. [ PubMed ] [ Google Scholar ]138. Baum AE, et al. Изследване за асоцииране в целия геном включва диацилглицерол киназа ета (DGKH) и няколко други гена в етиологията на биполярно разстройство. Мол. Психиатрия. 2008; 13 : 197–207. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]139. Манджи HK, Lenox RH. Сигнализация на протеин киназа С в мозъка: молекулярна трансдукция на стабилизация на настроението при лечение на маниакално-депресивни заболявания. Biol. Психиатрия. 1999; 46 : 1328–1351. [ PubMed ] [ Google Scholar ]140. Arnsten AFT, Manji HK. Мания: рационална невробиология. Бъдещ неврол. 2008; 3 : 125–131. [ Google Scholar ]141. Blumberg HP, et al. Възраст, ефекти на бързото колоездене и фармакотерапия върху вентралната префронтална кора при биполярно разстройство: проучване на напречното сечение. Biol. Психиатрия. 2006; 59 : 611–618. [ PubMed ] [ Google Scholar ]142. Bearden CE, et al. По-голяма кортикална плътност на сивото вещество при лекувани с литий пациенти с биполярно разстройство. Biol. Психиатрия. 2007; 62 : 7–16. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]143. Moore GJ, Bebchuk JM, Wilds IB, Chen G, Manji HK. Индуцирано от литий увеличение на човешкото мозъчно сиво вещество. The Lancet. 2000; 356 : 1241–1242. [ PubMed ] [ Google Scholar ]144. Bremner JD. Невроизобразяващи изследвания при посттравматично стресово разстройство. Curr. Психиатрия Rep.2002 ; 4 : 254–263. [ PubMed ] [ Google Scholar ]145. Southwick SM, et al. Роля на норадреналина в патофизиологията и лечението на посттравматично стресово разстройство. Biol. Психиатрия. 1999; 46 : 1192–1204. [ PubMed ] [ Google Scholar ]146. Morey RA, et al. Ролята на дистракторите, свързани с травма, върху невронните системи за обработка на работната памет и емоциите при посттравматично стресово разстройство. J. Psychiatr. Рез. 2009; 43 : 809–817. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]147. Wang L, et al. Префронталните механизми за изпълнителен контрол върху емоционалното разсейване се променят при голяма депресия. Психиатрия Res. 2008; 163 : 143–155. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]148. Хейуърд С, Санборн К. Пубертет и появата на половите различия в психопатологията. J. Adolesc. Здраве. 2002; 30 : 49–58. [ PubMed ] [ Google Scholar ]149. Shansky RM, et al. Естрогенът медиира половите различия при индуцирана от стреса префронтална дисфункция на кората. Мол. Психиатрия. 2004; 9 : 531–538. [ PubMed ] [ Google Scholar ]150. Jentsch JD, Roth RH, Taylor JR. Роля на допамина в поведенческите функции на префронталната кортикостриатална система: последици за психичните разстройства и психотропното лекарствено действие. Prog. Brain Res. 2000; 126 : 433–453. [ PubMed ] [ Google Scholar ]151. Markovac J, Goldstein GW. Пикомоларните концентрации на олово стимулират мозъчната протеинкиназа С. Природа. 1988; 334 : 71–73. [ PubMed ] [ Google Scholar ]152. Morgan RE, et al. Ранното излагане на олово води до трайни промени в продължително внимание, иницииране на реакция и реактивност на грешки. Невротоксикол. Тератол. 2001; 23 : 519–531. [ PubMed ] [ Google Scholar ]153. Cecil KM, et al. Намален обем на мозъка при възрастни с детска експозиция на олово. PLoS Med. 2008; 5 : e112. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]154. Невин Р. Как излагането на олово е свързано с временните промени в IQ, насилствените престъпления и неомъжената бременност. среда. Рез. 2000; 83 : 1–22. [ PubMed ] [ Google Scholar ]155. Wright JP, et al. Асоциация на пренатални и детски концентрации на олово в кръвта с престъпни арести в ранна възраст. PLoS Med. 2008; 5 : e101. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]156. Woolley DE. Перспектива на отравяне с олово в древността и настоящето. Невротоксикология. 1984; 5 : 353–361. [ PubMed ] [ Google Scholar ] Тази статия описва как падането на Римската империя вероятно включва отравяне с олово, тема от непосредствено значение за престъпността и неподходящото социално поведение в днешното общество.157. Brozoski T, Brown RM, Rosvold HE, Goldman PS. Когнитивен дефицит, причинен от регионално изчерпване на допамин в префронталната кора на маймуна резус. Наука. 1979; 205 : 929–931. [ PubMed ] [ Google Scholar ] Това е забележителен документ, който за първи път показа, че катехоламините са от съществено значение за дорзолатералните способности на префронталната работна памет. Въпреки че статията се фокусира върху допамина, сега е известно, че норадреналинът също има съществена роля.158. Bland ST, et al. Стресорната управляемост модулира индуцирания от стреса допамин и серотонинов излив и индуциран от морфин серотонинов излив в медиалната префронтална кора. Невропсихофармакология. 2003; 28 : 1589–1596. [ PubMed ] [ Google Scholar ]159. Clarke HF, Walker SC, Dalley JW, Robbins TW, Roberts AC. Когнитивната гъвкавост след префронтално изчерпване на серотонина е поведенчески и неврохимично специфична. Цереб. Кортекс. 2007; 17 : 18–27. [ PubMed ] [ Google Scholar ]160. Холмс А. Генетични вариации във функцията на серотонин на кортико-амигдала и риск от свързано със стреса заболяване. Невроски. Биобехав. Rev. 2008; 32 : 1293–1314. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]161. Williams GV, Rao SG, Goldman-Rakic PS. Физиологичната роля на 5-НТ 2А рецепторите в работната памет. J. Neurosci. 2002; 22 : 2843–2854. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]162. Boulougouris V, Glennon JC, Robbins TW. Дисоциируеми ефекти на селективните 5-HT 2A и 5-HT 2C рецепторни антагонисти върху серийното пространствено обръщане при плъхове Невропсихофармакология. 2008; 33 : 2007–2019. [ PubMed ] [ Google Scholar ]163. Голдман-Ракич П.С. В: Наръчник по физиология, Нервна система, Висши функции на мозъка. Plum F, редактор. Кн. V. Bethesda: Американско физиологично общество; 1987. с. 373–417. [ Google Scholar ] Тази класическа статия описва паралелните анатомични вериги, които стоят в основата на представителните знания.164. Прайс JL, Amaral DG. Авторадиографско изследване на проекциите на централното ядро на маймуната амигдала. J. Neurosci. 1981; 1 : 1242–1259. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]165. Цена JL, Carmichael ST, Drevets WC. Мрежи, свързани с орбиталната и медиалната префронтална кора; субстрат за емоционално поведение? Prog. Brain Res. 1996; 107 : 523–536. [ PubMed ] [ Google Scholar ]166. Ghashghaei HT, Barbas H. Пътища за емоции: взаимодействия на префронтални и предни темпорални пътища в амигдалата на маймуната резус. Неврология. 2002; 115 : 1261–1279. [ PubMed ] [ Google Scholar ]167. Simons JS, Henson RN, Gilbert SJ, Fletcher PC. Отделни форми на наблюдение на недвижими имоти от предната префронтална кора. J. Cogn. Невроски. 2008; 20 : 447–457. [ PMC безплатна статия ] [ PubMed ] [ Google Scholar ]168. Debiec J, LeDoux JE. Норадренергичното сигнализиране в амигдалата допринася за възстановяването на паметта на страха: последици от лечението за ПТСР. Ан. Ню Йорк акад. Sci. 2006; 1071 : 521–524. [ PubMed ] [ Google Scholar ]Професионално излагане на майката на изключително нискочестотни магнитни полета и риск от рак на мозъка при потомството. Li, Mclaughlin, et al. Причини и контрол на рака 20 (6): 945-55 (2009).Ракът причинява контрол
doi: 10.1007 / s10552-009-9311-5. Epub 2009 18 февруари.. 2009 август; 20 (6): 945-55.Професионално излагане на майката на изключително нискочестотни магнитни полета и риск от рак на мозъка при потомството
Принадлежности- PMID: 19224378
- DOI: 10.1007 / s10552-009-9311-5
Резюме
Цели: Да се изследва приноса на професионалното излагане на майката на изключително нискочестотни магнитни полета (ELF-MF) малко преди и по време на бременността върху честотата на детските мозъчни тумори.
Методи: В това проучване бяха включени общо 548 случая на инциденти и 760 здрави контроли, наети между 1980 и 2002 г. от две канадски провинции (Квебек и Онтарио) и бяха интервюирани техните майки. Количествената професионална експозиция на ELF-MF в единици microTesla беше оценена с помощта на индивидуални оценки на експозиция или матрица на експозиция на работа. Използвахме три показателя за анализ на експозицията: кумулативно, средно и достигнато максимално ниво.
Резултати: Използвайки средния показател на експозиция, измерен преди зачеването, се наблюдава повишен риск за астроглиални тумори (OR = 1,5, 95% CI = 1,0-2,4). По време на целия период на бременност се наблюдава значително повишен риск за астроглиални тумори, както и за всички детски мозъчни тумори със средна метрика (OR = 1,6, 95% CI = 1,1-2,5 и OR = 1,5, 95% CI = 1,1- 2.2, съответно). Въз основа на заглавията на длъжности се наблюдава двукратно увеличение на риска за астроглиални тумори (OR = 2,3, 95% CI = 0,8-6,3) и за всички детски мозъчни тумори (OR = 2,3, 95% CI = 1,0-5,4) сред операторите на шевни машини .
Заключения: Резултатите предполагат възможна връзка между професионалната експозиция на ELF-MF на майката и някои мозъчни тумори в тяхното потомство.
Подобни статии
- Професионално излагане на майката на изключително нискочестотни магнитни полета по време на бременност и детска левкемия.Епидемиология. 2003 юли; 14 (4): 437-41. doi: 10.1097 / 01.ede.0000078421.60231.bc.PMID: 12843769
- Професионално излагане на родители на изключително нискочестотни магнитни полета и детски рак: немско проучване за контрол на случаяAm J Epidemiol. 2010 г. 1 януари; 171 (1): 27-35. doi: 10.1093 / aje / kwp339. Epub 2009 25 ноември.PMID: 19942577
- Професионално излагане на изключително нискочестотни магнитни полета и рак на гърдата в постменопауза.Am J Ind Med. 2003 декември; 44 (6): 643-52. doi: 10.1002 / ajim.10264.PMID: 14635241
- [Канцерогенен риск от екстремно нискочестотни електромагнитни полета: състояние на техниката].Med Lav. 1994 ноември-декември; 85 (6): 447-62.PMID: 7731404 Преглед. Италиански.
- Сборен анализ на изключително нискочестотни магнитни полета и детски мозъчни тумори.Am J Epidemiol. 2010 1 октомври; 172 (7): 752-61. doi: 10.1093 / aje / kwq181. Epub 2010 9 август.PMID: 20696650 Безплатна статия от PMC. Преглед.
Цитирано от 6 статии
- Оценка на комбинирания ефект от изключително нискочестотно излагане на магнитно поле и оксидативен стрес върху метилирането на LINE-1 промотора в човешките нервни клетки.Radiat Environ Biophys. 2017 май; 56 (2): 193-200. doi: 10.1007 / s00411-017-0683-8. Epub 2017 3 март.PMID: 28258386
- Краткосрочното излагане на електромагнитно поле с изключително ниска честота увеличава броя на циркулиращите левкоцити и засяга сигнализирането на ос HPA при мишки.Биоелектромагнетика. 2016 г. октомври; 37 (7): 433-43. doi: 10.1002 / bem.21998. Epub 2016 24 август.PMID: 27553635 Безплатна статия от PMC.
- Оценка на професионалните рискове за изключително нискочестотни магнитни полета: Валидиране на емпиричен неекспертен подход.Предишна Med Rep.2016 от 31 май; 4: 148-54. doi: 10.1016 / j.pmedr.2016.05.017. eCollection 2016 дек.PMID: 27413676 Безплатна статия от PMC.
- Ефектът на нейонизиращото електромагнитно поле с честота 50 Hz в яйчниците на плъхове: Изследване с променлива електронна микроскопия.Int J Reprod Biomed. 2016 февруари; 14 (2): 125-32.PMID: 27200427 Безплатна статия от PMC.
- Генотоксичност, предизвикана от излагане на плода и бебето на магнитни полета и модулация на ефектите на йонизиращото лъчение.PLoS One. 2015 11 ноември; 10 (11): e0142259. doi: 10.1371 / journal.pone.0142259. eCollection 2015.PMID: 26559811 Безплатна статия от PMC.
Репродуктивни и развиващи ефекти на ЕМП в модели на гръбначни животни. Pourlis, AF Pathophysiology 16 (2-3): 179-89 (2009).
Въздействието на ЕМП върху възпроизводството и развитието върху модели на гръбначни животни
Принадлежности- PMID: 19272761
- DOI: 10.1016 / j.pathophys.2009.01.010
Резюме
Тази статия прави преглед на литературните данни за въздействието на електромагнитните полета (ЕМП), върху репродуктивните органи, както и върху пренаталното и постнаталното развитие на гръбначните животни. Бяха проучени статии за преглед, публикувани до 2001 г., относно репродуктивните и въздействията върху развитието на целия диапазон от честоти на електромагнитните полета. Експерименталните изследвания, публикувани от 2001 г. нататък, бяха обобщени. Специален фокус върху ефектите на радиочестотите, свързани с мобилната комуникация в гореспоменатите теми е направен. Според по-голямата част от разследванията не се получават силни ефекти по отношение на излагането на EMF на мобилната телефония при възпроизводството и развитието на животните.
Подобни статии
- Репродуктивни и тератологични ефекти на нискочестотните електромагнитни полета: преглед на in vivo и in vitro проучвания с животински модели.Тератология. 1999 април; 59 (4): 261-86. doi: 10.1002 / (SICI) 1096-9926 (199904) 59: 4 <261 :: AID-TERA12> 3.0.CO; 2-K.PMID: 10331529 Преглед.
- Противоракови ефекти върху плъхове Wistar, носещи лейомиосарком, след електромагнитно излъчване на резонансни радиочестоти.Hell J Nucl Med. 2007 май-август; 10 (2): 95-101.PMID: 17684584
- Електромагнитни полета (EMF): играят ли роля за здравето на околната среда на децата (CEH)?Int J Hyg Environment Health. 2007 г. октомври; 210 (5): 635-44. doi: 10.1016 / j.ijheh.2007.07.007. Epub 2007 31 август.PMID: 17765660 Преглед.
- Изключително нискочестотни електромагнитни полета като ефектори на клетъчни реакции in vitro: възможно активиране на имунните клетки.J Cell Biochem. 2004 1 септември; 93 (1): 83-92. doi: 10.1002 / jcb.20198.PMID: 15352165 Преглед.
- Изследване на краткосрочното излагане на електромагнитни полета върху репродуктивната способност на безгръбначните в полевата ситуация.Electromagn Biol Med. 2014 януари; 33 (1): 21-8. doi: 10.3109 / 15368378.2013.783846. Epub 2013 19 юни.PMID: 23781930
Цитирано от 8 статии
- Ефектът от пренаталното излагане на радиочестота от 2,4 GHz върху хистологията и експресията на остеокалцин и RUNX2 гена на предния край в NMRI мишка.J Lasers Med Sci. 2019 есен; 10 (4): 283-289. doi: 10.15171 / jlms.2019.46. Epub 2019 1 октомври.PMID: 31875120 Безплатна статия от PMC.
- Ефекти на фолиевата киселина върху бъбреците на плъхове, изложени на 900 MHz електромагнитно излъчване.J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 198-205. doi: 10.1016 / j.jmau.2017.06.001. Epub 2017 17 юни.PMID: 30023255 Безплатна статия от PMC.
- Преглед на електромагнитните полета (ЕМП) и репродуктивната система.Електронният лекар. 2016 юли 25; 8 (7): 2655-62. doi: 10.19082 / 2655. eCollection 2016 юли.PMID: 27648194 Безплатна статия от PMC. Преглед.
- Ефекти от 3 Hz и 60 Hz изключително нискочестотни електромагнитни полета върху подобно на тревожност поведение, запазване на паметта при пасивно избягване и електрофизиологични свойства на мъжките плъхове.J Lasers Med Sci. 2016 Пролет; 7 (2): 120-5. doi: 10.15171 / jlms.2016.20. Epub 2016 27 март.PMID: 27330708 Безплатна статия от PMC.
- Проучване на различни Samsung с интелигентни мобилни телефони Nokia в специфичното електрическо поле на главата.Glob J Health Sci. 2016 г. 1 септември; 8 (9): 53967. doi: 10.5539 / gjhs.v8n9p251.PMID: 27157169 Безплатна статия от PMC.
Пренатално и постнатално излагане на използването на мобилни телефони и поведенчески проблеми при деца. Divan, Kheifets, et al. Епидемиология19 (4): 523-29 (2008).
Пренатално и постнатално излагане на употреба на мобилен телефон и поведенчески проблеми при деца
Принадлежности- PMID: 18467962
- DOI: 10.1097 / EDE.0b013e318175dd47
Резюме
Контекст: Световната здравна организация подчерта необходимостта от проучване на възможните ефекти от радиочестотните полета при децата. Изследвахме връзката между пренаталното и постнаталното излагане на мобилни телефони и поведенческите проблеми при малки деца.
Методи: Майките са били назначени в датската национална кохорта за раждане в началото на бременността. Когато децата на тези бременности достигнат 7-годишна възраст през 2005 и 2006 г., майките са помолени да попълнят въпросник относно текущото здравословно състояние и поведенческия статус на децата, както и миналото излагане на употреба на мобилен телефон. Майките оцениха проблемите на поведението на детето, като използваха въпросника „Сила и трудности“.
Резултати: Майките на 13 159 деца попълниха последващия въпросник, отчитащ използването на мобилни телефони по време на бременност, както и текущата употреба на мобилни телефони от детето. Наблюдавани са по-големи коефициенти на шансове за поведенчески проблеми при деца, които са имали възможно пренатално или постнатално излагане на употреба на мобилен телефон. След корекция за потенциални смутители, съотношението на шансовете за по-висок резултат от общите поведенчески проблеми е 1,80 (95% доверителен интервал = 1,45-2,23) при деца с пренатална и постнатална експозиция на мобилни телефони.
Заключения: Излагането на мобилни телефони преди раждането и в по-малка степен след раждането е свързано с поведенчески затруднения като проблеми с емоционалността и хиперактивността около възрастта на влизане в училище. Тези асоциации може да са безпричинни и да се дължат на неизмеримо объркване. Ако са реални, те биха били от значение за общественото здраве предвид широкото използване на тази технология.
Коментирайте в
- Търсенето на въздействието върху околната среда върху здравето на децата: навигация между Scylla и Charybdis.Епидемиология. 2008 юли; 19 (4): 530-1. doi: 10.1097 / EDE.0b013e31817ae59d.PMID: 18497700
- Трябва ли епидемиолозите винаги да публикуват резултатите си? Да, почти винаги.Епидемиология. 2008 юли; 19 (4): 532-3. doi: 10.1097 / EDE.0b013e318177813d.PMID: 18497701
- Ниски предишни + плашещи последици = възпалителна епидемиология?Епидемиология. 2008 юли; 19 (4): 534-5. doi: 10.1097 / EDE.0b013e3181775e23.PMID: 18497702
- Използване на мобилен телефон на майката и поведенчески проблеми при деца.Епидемиология. 2009 март; 20 (2): 312. doi: 10.1097 / EDE.0b013e318188754d.PMID: 19234419 Няма наличен резюме.
Подобни статии
- Използване на мобилен телефон и поведенчески проблеми при малки деца.J Epidemiol Community Health. 2012 юни; 66 (6): 524-9. doi: 10.1136 / jech.2010.115402. Epub 2010 7 дек.PMID: 21138897
- Използване на мобилен телефон и безжичен телефон по време на бременност и проблеми с поведението при 5-годишни деца.J Epidemiol Community Health. 2013 май; 67 (5): 432-8. doi: 10.1136 / jech-2012-201792. Epub 2013 5 февруари.PMID: 23386674
- Пренатално и постнатално излагане на тютюн и поведенчески проблеми при 10-годишни деца: резултати от проучването на GINI-plus за проспективно раждане.Environmental Health Perspect. 2010 януари; 118 (1): 150-4. doi: 10.1289 / ehp.0901209.PMID: 20056582 Безплатна статия от PMC.
- Пренатален тютюнев дим и постнатално излагане на вторичен дим и детско невроразвитие.Curr Opin Pediatr. 2008 април; 20 (2): 184-90. doi: 10.1097 / MOP.0b013e3282f56165.PMID: 18332716 Преглед.
- Използване на мобилен телефон и експозиция при деца.Биоелектромагнетика. 2005; Допълнение 7: S45-50. doi: 10.1002 / bem.20129.PMID: 16142783 Преглед.
Цитирано от 25 статии
- Обогатената околна среда намалява когнитивното увреждане при възрастни плъхове с пренатална експозиция на мобилен телефон.Невроски от предно стареене. 2020 юни 4; 12: 162. doi: 10.3389 / fnagi.2020.00162. eCollection 2020.PMID: 32581772 Безплатна статия от PMC.
- Ранното излагане на импулсни LTE радиочестотни полета причинява трайни промени в активността и поведението на мишки C57BL / 6 J.Биоелектромагнетика. 2019 октомври; 40 (7): 498-511. doi: 10.1002 / bem.22217. Epub 2019 15 септември.PMID: 31522469 Безплатна статия от PMC.
- Рискове за здравето и благосъстоянието от радиочестотното излъчване, излъчвано от клетъчни телефони и други безжични устройства.Front Public Health. 2019 август 13; 7: 223. doi: 10.3389 / fpubh.2019.00223. eCollection 2019.PMID: 31457001 Безплатна статия от PMC. Преглед.
- Асоциации на използването на мобилен телефон на майката по време на бременност с продължителност на бременността и растеж на плода в 4 кохорти при раждане.Am J Epidemiol. 2019 юли 1; 188 (7): 1270-1280. doi: 10.1093 / aje / kwz092.PMID: 30995291 Безплатна статия от PMC.
- Промени в профила на хипокампалния липидом и транскриптома, предизвикани от остро излагане на мишки на облъчване на мобилен телефон GSM 1800 MHz: Изследователско проучванеМозък Behav. 2018 юни; 8 (6): e01001. doi: 10.1002 / brb3.1001. Epub 2018 22 май.PMID: 29786969 Безплатна статия от PMC.
Ефекти от пренаталното излагане на 900 MHz електромагнитно поле върху зъбната извивка на плъхове: Стереологично и хистопатологично проучване. Odaci, et al. Brain Research 1238: 224–229
Ефекти от пренаталното излагане на 900 MHz електромагнитно поле върху зъбната извивка на плъхове: стереологично и хистопатологично проучване
Принадлежности- PMID: 18761003
- DOI: 10.1016 / j.brainres.2008.08.013
Резюме
Електромагнитните полета (ЕМП) инхибират образуването и диференциацията на нервните стволови клетки по време на ембрионалното развитие. В това проучване бяха изследвани ефектите от пренаталното излагане на ЕМП върху броя на грануларните клетки в зъбната извивка на 4-седмични плъхове. Този експеримент използва контролна (Cont) група и група, изложена на EMF (EMF) (по три бременни плъхове всяка група). Групата с ЕМП се състоеше от шест потомци (n = 6) от бременни плъхове, които бяха изложени на ЕМП до 900 мегахерца (MHz) за 60 минути на ден между първия и последния ден на бременността. Контролната група се състоеше от пет потомци (n = 5) от бременни плъхове, които изобщо не бяха лекувани. Потомството е било жертвано, когато е било на 4 седмици. Броят на грануларните клетки в зъбната извивка беше анализиран с помощта на оптичната фракционираща техника. Резултатите показват, че пренаталната експозиция на ЕМП води до намаляване на броя на грануларните клетки в зъбната извивка на плъховете (P <0,01). Това предполага, че пренаталната експозиция на 900 MHz EMF влияе върху развитието на гранулатните клетки на зъбните извивки в хипокампуса на плъхове. Загубата на клетки може да бъде причинена от инхибиране на гранулозно-клетъчната неврогенеза в зъбния извив.
Подобни статии
- Излагането на електромагнитно поле от 900 MHz засяга качествените и количествените характеристики на хипокампалните пирамидални клетки при възрастни женски плъхове.Brain Res. 2009 г., 10 април; 1265: 178-85. doi: 10.1016 / j.brainres.2009.02.011. Epub 2009 20 февруари.PMID: 19230827
- Ефект от пренаталното излагане на противовъзпалително лекарство върху броя на невроните в роговия амонис и зъбната извивка на хипокампуса на плъхове: стереологично проучване.Brain Res. 2007 г., 5 януари; 1127 (1): 185-92. doi: 10.1016 / j.brainres.2006.10.026. Epub 2006 21 ноември.PMID: 17123485
- Хроничното пренатално излагане на 900 мегагерцово електромагнитно поле предизвиква пирамидална клетъчна загуба в хипокампуса на новородени плъхове.Toxicol Ind Health. 2009 юли; 25 (6): 377-84. doi: 10.1177 / 0748233709106442.PMID: 19671630
- Морфологично развитие и съзряване на гранулирани невронови дендрити в зъбната извивка на плъх.Prog Brain Res. 2007; 163: 167-81. doi: 10.1016 / S0079-6123 (07) 63010-6.PMID: 17765718 Преглед.
- Невронната първична ресничка: двигател на неврогенезата и формирането на паметта в хипокампалния зъбен извивка?Клетъчен сигнал. 2009 септември; 21 (9): 1351-5. doi: 10.1016 / j.cellsig.2009.02.013. Epub 2009 26 февруари.PMID: 19249355 Преглед.
Цитирано от 20 статии
- Излагането на електромагнитно поле по време на бременността влияе неблагоприятно на електрофизиологичните свойства на клетките от Purkinje при потомство на плъхове.J Biomed Phys Eng. 2020 г. 1 август; 10 (4): 433-440. doi: 10.31661 / jbpe.v0i0.560. eCollection 2020 августPMID: 32802791 Безплатна статия от PMC.
- Обогатената околна среда намалява когнитивното увреждане при възрастни плъхове с пренатална експозиция на мобилен телефон.Невроски от предно стареене. 2020 юни 4; 12: 162. doi: 10.3389 / fnagi.2020.00162. eCollection 2020.PMID: 32581772 Безплатна статия от PMC.
- Излагането на радиочестота на клетъчния телефон променя нивата на кортикотропинов хормон и хистологията на мозъка и надбъбречните жлези при мъжки плъх Wistar.Iran J Basic Med Sci. 2018 декември; 21 (12): 1269-1274. doi: 10.22038 / ijbms.2018.29567.7133.PMID: 30627371 Безплатна статия от PMC.
- Радиации и мъжка плодовитост.Reprod Biol Endocrinol. 2018 декември 9; 16 (1): 118. doi: 10.1186 / s12958-018-0431-1.PMID: 30445985 Безплатна статия от PMC. Преглед.
- Защитни ефекти на мелатонин и омега-3 върху хипокампуса и малкия мозък на възрастни плъхове албиноси Wistar, изложени на електромагнитни полета.J Microsc Ultrastruct. 2017 октомври-декември; 5 (4): 230-241. doi: 10.1016 / j.jmau.2017.05.006. Epub 2017 1 юни.PMID: 30023259 Безплатна статия от PMC.
(2008).
Излагането на радиация на мобилен телефон нагоре регулира гените на апоптоза в първичните култури на неврони и астроцити. Zhao, et al. Science Digest 412: 34–38 (2007).
Излагането на радиация на клетъчния телефон нагоре регулира гените на апоптоза в първичните култури на неврони и астроцити
Принадлежности- PMID: 17187929
- PMCID: PMC2713174
- DOI: 10.1016 / j.neulet.2006.09.092
Резюме
Ефектите върху здравето на излагането на радиация на мобилни телефони са нарастваща тревога на обществеността. Това проучване изследва дали експресията на гени, свързани с пътищата на клетъчната смърт, не е регулирана в първично култивирани неврони и астроцити чрез излагане на работещ мобилен телефон на Глобалната система за мобилна комуникация (GSM) с честота 1900MHz. Първичните култури бяха изложени на емисии от клетъчни телефони в продължение на 2 часа. Използвахме масивен анализ и RT-PCR в реално време, за да покажем регулация на експресията на гена на каспаза-2, каспаза-6 и Asc (свързан с апоптоза спек-подобен протеин, съдържащ карта) в неврони и астроцити. Регулирането нагоре се наблюдава както в режим "включен", така и в режим на готовност в невроните, но само в режим "включен" в астроцитите. Освен това астроцитите показаха регулиране на гена Bax. Ефектите са специфични, тъй като не се наблюдава повишена регулация за други гени, свързани с апоптоза, като каспаза-9 нито в невроните, нито в астроцитите, или Bax в невроните. Резултатите показват, че дори относително краткотрайно излагане на радиочестотни емисии от клетъчния телефон може да регулира нагоре елементи на апоптотични пътища в клетки, получени от мозъка, и че невроните изглежда са по-чувствителни към този ефект от астроцитите.
Фигури


Подобни статии
- Електромагнитните полета засягат нивата на транскрипт на гените, свързани с апоптозата, в невронни клетки-предшественици на ембрионални стволови клетки.FASEB J. 2005 октомври; 19 (12): 1686-8. doi: 10.1096 / fj.04-3549fje. Epub 2005 22 август.PMID: 16116041
- Доказателства за участие на Bax и p53, но не и на каспази, в радиационно-индуцирана клетъчна смърт на култивирани постнатални хипокампални неврони.J Neurosci Res. 1998 г. 15 декември; 54 (6): 721-33. doi: 10.1002 / (SICI) 1097-4547 (19981215) 54: 6 <721 :: AID-JNR1> 3.0.CO; 2-1.PMID: 9856857
- Апоптозата се сигнализира рано от ниски дози йонизиращо лъчение в радиационно-индуциран страничен ефект.Mutat Res. 2013 януари-февруари; 741-742: 35-43. doi: 10.1016 / j.mrfmmm.2013.02.001. Epub 2013 20 февруари.PMID: 23454491
- Биохимични модификации и увреждане на невроните в мозъка на млади и възрастни плъхове след продължително излагане на излъчване на мобилен телефон.Cell Biochem Biophys. 2014 ноември; 70 (2): 845-55. doi: 10.1007 / s12013-014-9990-8.PMID: 24801773
- Индуцирана от химиотерапия клетъчна смърт в първичните церебеларни гранули неврони, но не и в астроцитите: in vitro парадигма на диференциалната невротоксичност.J Neurochem. 2004 декември; 91 (5): 1067-74. doi: 10.1111 / j.1471-4159.2004.02774.x.PMID: 15569250
Цитирано от 28 статии
- Радиочестотни електромагнитни радиационни промени в поведението и тяхната възможна основа.Environment Sci Pollut Res Int. 2019 октомври; 26 (30): 30693-30710. doi: 10.1007 / s11356-019-06278-5. Epub 2019 28 август.PMID: 31463749 Преглед.
- Значителна зависимост на клетъчната жизнеспособност от експозицията на времето при ELF-EMF и RF-EMF In vitro проучвания.Int J Environment Res Public Health. 2019 юни 13; 16 (12): 2085. doi: 10.3390 / ijerph16122085.PMID: 31200435 Безплатна статия от PMC.
- Трансдермални пластири, заредени с L-цистеин HCL като стратегия за защита от мобилни телефони, излъчващи опасности от електромагнитно излъчване.Saudi Pharm J. 2019 г. януари; 27 (1): 112-125. doi: 10.1016 / j.jsps.2018.09.004. Epub 2018 5 септември.PMID: 30662314 Безплатна статия от PMC.
- Възможни ефекти от излагането на радиочестотно електромагнитно поле върху централната нервна система.Biomol Ther (Сеул). 2019 май 1; 27 (3): 265-275. doi: 10.4062 / biomolther.2018.152.PMID: 30481957 Безплатна статия от PMC. Преглед.
- Излагането на електромагнитна радиация 2450 MHz причинява дефицит на познание с митохондриална дисфункция и активиране на вътрешния път на апоптоза при плъхове.J Biosci. 2018 юни; 43 (2): 263-276.PMID: 29872015
Клетъчна смърт, предизвикана от GSM 900-MHz и DCS 1800-MHz излъчване на мобилна телефония. Panagopoulos, et al. Мутационни изследвания626, 69–78 (2006).
Клетъчна смърт, предизвикана от GSM 900-MHz и DCS 1800-MHz излъчване на мобилната телефония
Принадлежности- PMID: 17045516
- DOI: 10.1016 / j.mrgentox.2006.08.008
Резюме
В настоящото изследване се използва TUNEL (терминален дезоксинуклеотид трансфераза dUTP Nick End Labeling) - добре позната техника, широко използвана за откриване на фрагментирана ДНК в различни видове клетки - за откриване на клетъчна смърт (фрагментация на ДНК) в биологичен модел , ранните и средните етапи на оогенезата на насекомото Drosophila melanogaster. Мухите са били изложени in vivo на GSM 900-MHz (Глобална система за мобилни телекомуникации) или DCS 1800-MHz (Digital Cellular System) облъчване от общ цифров мобилен телефон, за няколко минути на ден през първите 6 дни от възрастния живот. Условията на експозиция бяха подобни на тези, на които е изложен потребител на мобилен телефон, и бяха определени според предишни наши проучвания [DJ Panagopoulos, A. Karabarbounis, LH Margaritis, Ефект на GSM 900-MHz излъчване на мобилен телефон върху репродуктивния капацитет на D. melanogaster, Electromagn. Biol. Med. 23 (1) (2004) 29-43; DJ Panagopoulos, N. Messini, A. Karabarbounis, AL Philippetis, LH Margaritis, Радиочестотното електромагнитно излъчване в рамките на "нивата на безопасност" променя физиологичната функция на насекомите, в: P. Kostarakis, P. Stavroulakis (Eds.), Proceedings of the Международен семинар на хилядолетието за биологичните ефекти на електромагнитните полета, Ираклион, Крит, Гърция, 17-20 октомври 2000 г., стр. 169-175, ISBN: 960-86733-0-5; DJ Panagopoulos, LH Margaritis, Ефекти на електромагнитните полета върху репродуктивната способност на D. melanogaster, в: P. Stavroulakis (Ed.), Biological Effects of Electromagnetic Fields, Springer, 2003, pp. 545-578], което беше показало голям спад в яйцекласата на същото насекомо, причинено от GSM лъчение. Нашите настоящи резултати предполагат, че намаляването на яйцекласването, съобщено по-рано, се дължи на дегенерация на голям брой яйцеклетки след фрагментация на ДНК на техните съставни клетки, индуцирана от двата вида радиация на мобилната телефония. Индуцирана клетъчна смърт се регистрира за първи път във всички видове клетки, съставляващи яйцеклетка (фоликуларни клетки, клетки на кърмачки и ооцит) и във всички етапи на ранния и средния оогенезис, от гермария до етап 10, по време на който се програмира клетъчната смърт не настъпва физиологично. Установено е, че гермариумът и фазите 7-8 са най-чувствителните етапи на развитие също в отговор на електромагнитното напрежение, индуцирано от GSM и DCS полетата и, освен това,
Подобни статии
- Сравнение на биологичните ефекти между непрекъснато и периодично излагане на излъчване на мобилен телефон GSM-900 MHz: Откриване на апоптотични характеристики на клетъчна смъртMutat Res. 2010 юли 19; 700 (1-2): 51-61. doi: 10.1016 / j.mrgentox.2010.05.008. Epub 2010 21 май.PMID: 20472095
- Сравнение на биоактивността между GSM 900 MHz и DCS 1800 MHz излъчване на мобилна телефония.Electromagn Biol Med. 2007; 26 (1): 33-44. doi: 10.1080 / 15368370701205644.PMID: 17454081
- Ефектът на продължителността на експозицията върху биологичната активност на радиацията на мобилната телефония.Mutat Res. 2010 юни 17; 699 (1-2): 17-22. doi: 10.1016 / j.mrgentox.2010.04.010. Epub 2010 23 април.PMID: 20399887
- Мета-анализ за невро-поведенчески ефекти поради излагане на електромагнитно поле, излъчвано от GSM мобилни телефони.Occup Environ Med. 2008 май; 65 (5): 342-6. doi: 10.1136 / oem.2006.031450. Epub 2007 10 октомври.PMID: 17928386 Преглед.
- Електромагнитни полета (EMF): играят ли роля за здравето на околната среда на децата (CEH)?Int J Hyg Environment Health. 2007 г. октомври; 210 (5): 635-44. doi: 10.1016 / j.ijheh.2007.07.007. Epub 2007 31 август.
Свръхвисокочестотното електромагнитно поле облъчване по време на бременност води до увеличаване на честотата на микроядрите на еритроцитите при потомството на плъхове. Ferreira, Knakievicz, et al. Науки за живота 80 (1): 43-50 (2006).
Излъчването на ултрависокочестотно електромагнитно поле по време на бременност води до увеличаване на честотата на микроядрата на еритроцитите при потомството на плъхове
Принадлежности- PMID: 16978664
- DOI: 10.1016 / j.lfs.2006.08.018
Резюме
Мобилните телефони и техните базови станции са важен източник на ултрависокочестотно електромагнитно поле (UHF-EMF) и тяхното използване се увеличава навсякъде по света. Епидемиологичните проучвания показват, че нискоенергийният UHF-EMF, излъчен от клетъчния телефон, може да причини биологични ефекти, като увреждане на ДНК и промени в окислителния метаболизъм. Цитогенетичен тест in vivo за бозайници, анализът на микроядрата (MN), се използва за изследване на появата на хромозомни увреждания в еритроцитите от потомство на плъхове, изложени на нетермичен UHF-EMF от клетъчен телефон по време на тяхната ембриогенеза; облъчената група показа значително увеличение на появата на MN. За да се изследва дали UHF-EMF може също така да промени окислителните параметри в периферната кръв и в черния дроб - важна хематопоетична тъкан при ембриони и новородени от плъхове - ние също измерихме активността на антиоксидантните ензими, количествено определили общото съдържание на сулфхидрил, протеиновите карбонилни групи, реагиращи на тиобарбитурова киселина видове и пълна неензимна антиоксидантна защита. Не са открити значителни разлики в който и да е окислителен параметър на кръвта и черния дроб на потомството. Средният брой на малките във всяко котило също не е променен значително. Нашите резултати показват, че при нашите експериментални условия UHF-EMF е в състояние да индуцира генотоксичен отговор в хематопоетичната тъкан по време на ембриогенезата чрез неизвестен механизъм. протеинови карбонилни групи, реагиращи на тиобарбитурова киселина видове и обща неензимна антиоксидантна защита. Не са открити значителни разлики в който и да е окислителен параметър на кръвта и черния дроб на потомството. Средният брой на малките във всяко котило също не е променен значително. Нашите резултати показват, че при нашите експериментални условия UHF-EMF е в състояние да индуцира генотоксичен отговор в хематопоетичната тъкан по време на ембриогенезата чрез неизвестен механизъм. протеинови карбонилни групи, реагиращи на тиобарбитурова киселина видове и обща неензимна антиоксидантна защита Не са открити значителни разлики в който и да е окислителен параметър на кръвта и черния дроб на потомството. Средният брой на малките във всяко котило също не е променен значително. Нашите резултати показват, че при нашите експериментални условия UHF-EMF е в състояние да индуцира генотоксичен отговор в хематопоетичната тъкан по време на ембриогенезата чрез неизвестен механизъм.
Подобни статии
- Ефекти на оксидативен стрес върху централната нервна система на плъхове след остро излагане на ултрависокочестотни електромагнитни полета.Биоелектромагнетика. 2006 септември; 27 (6): 487-93. doi: 10.1002 / bem.20233.PMID: 16715528
- Ефект на 910-MHz електромагнитно поле върху костния мозък на плъх.ScientificWorldJournal. 2004 г. 20 октомври; 4 Suppl 2: 48-54. doi: 10.1100 / tsw.2004.178.PMID: 15517102 Безплатна статия от PMC.
- Оценка на цитогенотоксичните увреждания при незрели и зрели плъхове, изложени на 900 MHz радиочестотни електромагнитни полета.Int J Radiat Biol. 2013 ноември; 89 (11): 985-92. doi: 10.3109 / 09553002.2013.809170. Epub 2013 21 юни.PMID: 23718180
- Излагане на ниско ниво на радиочестотни електромагнитни полета: ефекти върху здравето и нужди от изследвания.Биоелектромагнетика. 1998; 19 (1): 1-19.PMID: 9453702 Преглед.
- Изключително нискочестотни електромагнитни полета като ефектори на клетъчни реакции in vitro: възможно активиране на имунните клетки.J Cell Biochem. 2004 1 септември; 93 (1): 83-92. doi: 10.1002 / jcb.20198.PMID: 15352165 Преглед.
Цитирано от 4 статии
- Реални спрямо симулирани експозиции на мобилни телефони в експериментални изследвания.Biomed Res Int. 2015; 2015: 607053. doi: 10.1155 / 2015/607053. Epub 2015 5 август.PMID: 26346766 Безплатна статия от PMC. Преглед.
- Мобилни телефони и тумори на главата. Несъответствията в причинно-следствените връзки в епидемиологичните проучвания - как възникват?Environment Health. 2011 юни 17; 10: 59. doi: 10.1186 / 1476-069X-10-59.PMID: 21679472 Безплатна статия от PMC.
- Изследвания за увреждане на ДНК и честота на микроядра при професионална експозиция на електромагнитни полета (EMF), излъчвани от терминали за видео дисплей (VDT).Genet Mol Biol. 2010 януари; 33 (1): 154-8. doi: 10.1590 / S1415-47572010005000010. Epub 2010 1 март.PMID: 21637620 Безплатна статия от PMC.
- Отговор на коментари на A. Lerchl относно "Радиочестотните електромагнитни полета (UMTS, 1,950 MHz) предизвикват генотоксични ефекти in vitro при човешки фибробласти, но не и в лимфоцити", публикуван от C. Schwarz et al. 2008 г.Int Arch Occup Environment Health. 2009 януари; 82 (2): 279-83. doi: 10.1007 / s00420-008-0330-4. Epub 2008 15 май.PMID: 18481076
Синдром на дефицит на вниманието и хиперактивност. Biederman, J. & Faraone, SV Lancet 366, 237–248 (2005).
Синдром на дефицит на вниманието и хиперактивност
Принадлежности- PMID: 16023516
- DOI: 10.1016 / S0140-6736 (05) 66915-2
Грешка в
- Лансет. 2006 г., 21 януари; 367 (9506): 210
Резюме
Разстройство с хиперактивност с дефицит на вниманието (ADHD) е разстройство на невнимание, импулсивност и хиперактивност, което засяга 8-12% от децата по света. Въпреки че процентът на ADHD пада с възрастта, поне половината от децата с това разстройство ще имат влошаващи симптоми в зряла възраст. Изследванията на близнаци, осиновяване и молекулярни генетични изследвания показват, че ADHD е силно наследствен, а други открития са регистрирали акушерски усложнения и психосоциални неприятности като предразполагащи рискови фактори. Сближаващите се данни от проучвания върху животни и хора предполагат нарушаване на регулирането на фронтално-подкорко-мозъчните катехоламинергични вериги в патофизиологията на ADHD, а изследванията на молекулярните образи показват, че аномалиите на допаминовия транспортер водят до нарушена невротрансмисия. Изследвания през последното десетилетие показаха безопасността и ефективността на новите нестимулиращи лекарства и дългодействащи формулировки на метилфенидат и амфетамин. Други разследвания също изясниха подходящата роля на целенасочените психосоциални лечения в контекста на продължаващата фармакотерапия.
Подобни статии
- Взаимодействащи ефекти на гена на допаминов транспортер и психосоциални неблагополучия върху симптомите на дефицит на внимание / хиперактивност при 15-годишни от високорискова проба в общносттаАрх генерал психиатрия. 2007 май; 64 (5): 585-90. doi: 10.1001 / archpsyc.64.5.585.PMID: 17485610
- Дефицит на внимание / хиперактивност: селективен преглед.Психиатрия на Biol. 2005 1 юни; 57 (11): 1215-20. doi: 10.1016 / j.biopsych.2004.10.020. Epub 2004 18 дек.PMID: 15949990 Преглед.
- Преглед на патофизиологията, етиологията и лечението на разстройство с хиперактивност с дефицит на вниманието (ADHD).Ann Pharmacother. 2014 февруари; 48 (2): 209-25. doi: 10.1177 / 1060028013510699. Epub 2013 1 ноември.PMID: 24259638 Преглед.
- Дефицит на внимание / хиперактивност и допаминергичните хипотези.Експерт Rev Neurother. 2010 април; 10 (4): 587-601. doi: 10.1586 / ern.10.17.PMID: 20367210 Преглед.
- Предшестващо състояние, клинични особености и лечение на разстройство с хиперактивност с дефицит на внимание при деца.Експерт Opin Pharmacother. 2013 г. октомври; 14 (14): 1885-906. doi: 10.1517 / 14656566.2013.818977. Epub 2013 19 юли.PMID: 23865438 Преглед.
Цитирано от 417 статии
- Дефицит на внимание / хиперактивност с разстройство Пациентите може да са недиагностицирани нарколепсия.Кюреус. 2020 юни 4; 12 (6): e8436. doi: 10.7759 / cureus.8436.PMID: 32642351 Безплатна статия от PMC.
- Мигрена, разстройство с хиперактивност с дефицит на вниманието и време на екрана при деца, посещаващи институция за третична грижа в Шри Ланка: свързани ли са те?BMC Neurol. 2020 г. 8 юли; 20 (1): 275. doi: 10.1186 / s12883-020-01855-5.PMID: 32640997 Безплатна статия от PMC.
- Тенденции в психиатричните диагнози, лекарствата и психологичните терапии в голям шведски регион: проучване, основано на населението.BMC Психиатрия. 2020 г., 23 юни; 20 (1): 328. doi: 10.1186 / s12888-020-02749-z.PMID: 32576173 Безплатна статия от PMC.
- Намалява ли медикаментите за разстройство с хиперактивност с дефицит на вниманието предприемачеството?Health Econ. 2020 г. септември; 29 (9): 1071-1077. doi: 10.1002 / hec.4120. Epub 2020 22 юни.PMID: 32567214 Безплатна статия от PMC.
- Присъствие на спешно отделение за нараняване и поведение, предполагащи разстройство с хиперактивност с дефицит на вниманието (ADHD): проучване в напречно сечение.BMC Педиатър. 2020 г., 29 май; 20 (1): 266. doi: 10.1186 / s12887-020-02166-x.PMID: 32471498 Безплатна статия от PMC.
Разстройство с дефицит на внимание / хиперактивност: преглед на етиологията и преглед на литературата, свързана с корелатите и резултатите от жизнения път на мъжете и жените Brassett-Harknett, A. & Butler, N. Clinical Psychology Review 27,188–210 (2005).
Дефицит на внимание / хиперактивност: Преглед на етиологията и преглед на литературата, свързана с корелатите и резултатите от жизнения цикъл на мъжете и жените















II. Ефекти върху малките деца
Електромагнитни полета, импулсно радиочестотно излъчване и епигенетика: Как безжичните технологии могат да повлияят на развитието на детството. Sage, C. & Burgio, E. Развитие на детето (2017).
Проспективен кохортен анализ на употребата на мобилни телефони и емоционални и поведенчески трудности при деца. Судан, М, et al. Списание за епидемиология и здраве на общността (2016).
Защо децата поглъщат повече микровълнова радиация от възрастните: Последиците. Morgan, Kesari, et al. Вестник по микроскопия и ултраструктура 2 (4): 196-204 (2014).
Епидемиологични характеристики на собствеността и използването на мобилни телефони при корейски деца и юноши. Byun, Yoon-Hwan, et al. Екологично здраве и токсикология 28 (2013).
Проспективно проучване на вътреутробната експозиция на магнитни полета и риска от детско затлъстяване. Li, De-Kun, et al. Научни доклади 2.540 (2012).
Излагане на изключително нискочестотни магнитни полета и риск от рак на детството: Актуализация на епидемиологичните доказателства. Шюц и Йоаким. Напредък в биофизиката и молекулярната биология 107 (3): 339-42 (2011).
Употреба на мобилен телефон и поведенчески проблеми при малки деца. Divan, Kheifets, et al. Journal of Epidemiol Community Health 66 (6): 524-9 (2010).
Мобилни телефони, радиочестотни полета и ефекти върху здравето при деца-епидемиологични изследвания. Фейхтинг, Мария. Напредък в биофизиката и молекулярната биология 107 (3): 343-348 (2010).
Излагане на радиочестотни електромагнитни полета и поведенчески проблеми при баварски деца и юноши. Thomas, Silke, et al. Европейско списание за епидемиология 25 (2): 135-41 (2009).
Чувствителността на децата към електромагнитни полета. Repacholi, et al. Девентър. Journal of Pediatrics 116 (2): 303-313 (2005).
III. Мозъчни тумори
Симулация на честотата на злокачествени мозъчни тумори в кохорти на раждането, които започнаха да използват мобилни телефони, когато за първи път станаха популярни в Япония. Sato, Y., Kojimahara, N. и Yamaguchi, N. Биоелектромагнетика 40 (3): 143-149 (2019).
Доклад за крайните резултати по отношение на мозъчни и сърдечни тумори при плъхове Sprague-Dawley, изложени от пренатален живот Unitl Естествена смърт до поле за радиочестота на мобилен телефон Представител на 1,8 GHz GSM базова станция Емисия на околната среда. Falcioni, L, et al. Изследвания на околната среда (2018).
Излагането на радиочестотни промени на клетъчния телефон Нива на кортикотропинов хормон и хистология на мозъка и надбъбречните жлези при мъжки плъх Wistar. Shahabi, S., et al. Ирански списание за основни медицински науки 21: 1269-1274 (2018).
Мозъчни тумори: Повишаване на мултиформената честота на глиобластома в Англия 1995-2015 предполага неблагоприятен екологичен фактор или начин на живот Philips, A., et al. Вестник за околната среда и общественото здраве (2018).
Радиочестотното излъчване от 2100 MHz на 3G-мобилен телефон и окислителните щети на ДНК в мозъка. Sahin, Ozgur, et al. Journal of Chemical Neuroanatomy 75 (B): 94-98 (2016).
Употреба на мобилни телефони и безжични телефони и риск за глиома - анализ на обединени проучвания за контрол на случаите в Швеция 1997-2003 и 2007-2009. Хардел и Карлберг. Патофизиология 22 (1): 1-13 (2015).
Излъчването на мобилен телефон причинява мозъчни тумори и трябва да бъде класифицирано като вероятен канцероген за човека. Morgan, Miller, et al. Международен вестник по онкология 46: 1865-1871 (2015).
Използване на мобилни телефони и мозъчни тумори в проучването CERENAT Case-Control. Coureau, Bouvier, et al. Медицина на труда и околната среда 71 (7): 514-22 (2014).
Използването на мобилни телефони и безжични телефони е свързано с повишен риск за глиома и акустична неврома. Hardell, Carberg, et al. Патофизиология 20 (2): 85-110 (2013).
Мобилни телефони и тумори на главата: критичен анализ на епидемиологични проучвания за контрол на случаите. Levis, Minicuci, et al. Отворени науки за околната среда 6 (1): 1-12 (2012).
За връзката между глиома, безжични телефони, наследственост и йонизиращо лъчение. Карлберг и Хардел. PathoPhysiology19 (4): 243-252 (2012).
Мобилни телефони и тумори на главата. Несъответствията в причинно-следствените връзки в епидемиологичните проучвания - как възникват? Levis, Minicuci, et al. Екологично здраве 10:59 (2011).
Индикации за възможен риск от мозъчен тумор при изследвания на мобилни телефони: Трябва ли да се тревожим? Кардис и Садецки. Медицина на труда и околната среда 68: 169-171 (2011).
Оценка на риска от мозъчни тумори от използването на мобилен телефон: Публикувани проучвания за контрол на случая. Морган, LL. Патофизиология 16 (2-3): 137-147 (2009).
Клетъчни телефони и мозъчни тумори: Преглед, включващ дългосрочните епидемиологични данни. Khurana, Teo, et al. Хирургична неврология72 (3): 205-14 (2009).
Епидемиологични доказателства за връзка между използването на безжични телефони и туморни заболявания. Hardell, Carlberg, et al. Патофизиология 16 (2-3): 113-122 (2009).
Хистопатологични изследвания на мозъци на плъхове след продължително излагане на излъчване на мобилен телефон GSM. Grafström, Gustav, et al. Бюлетин за мозъчни изследвания 77 (5): 257-63 (2008).
Използване на мобилен телефон и риск от акустична неврома. Lonn, Ahlbom, et al. Епидемиология 15 (6): 653-659 (2004).
IV. Тумори на паротидната жлеза
Влияние на ръчни мобилни телефони върху паротида: Кохортно проучване. Ranjitha, G., et al. Вестник на Индийската академия по орална медицина и радиология 29: 254-258 (2017).
Увеличава ли използването на мобилен телефон шансовете за развитие на тумор на паротидната жлеза? Систематичен преглед и мета-анализ. De Siqueira, de Souza, et al. Вестник по орална патология и медицина 45 (11) (2016).
Сборен анализ на проучвания за контрол на случаите на диагностицирани акустични невроми 1997-2003 и 2007-2009 и използване на мобилни и безжични телефони. Hardell, Carlberg, et al. Международен вестник по онкология 43 (4): 1036-144 (2015).
Използване на възгледите на Hill от 1965 г. за оценка на силните страни на доказателствата за риска за мозъчни тумори, свързани с използването на мобилни и безжични телефони. Хардел и Карлберг. Прегледи за здравето на околната среда 28 (2-3): 97-106 (2013).
Case-Control проучване на използването на мобилни и безжични телефони и риска от злокачествен меланом в областта на главата и шията. Hardell, Carlberg, et al. Патофизиология 18 (4): 325-333 (2011).
Връзка между употребата на клетъчен телефон и злокачествените образувания на епителната паротидна жлеза. Duan, Zhang, et al. Глава на клиничната хартия и онкология 40 (9): 966-7 (2011).
Използване на мобилни телефони и риск от тумори: мета-анализ. Mynf, Ju, et al. Вестник по клинична онкология 27 (33): 5565-72 (2009).
Мобилен телефон, безжични телефони и рискът от мозъчни тумори. Хардел и Карлберг. Международен вестник по онкология 35 (1): 5-17 (2009).
Последици за общественото здраве на безжичните технологии. Мъдрец и Дърводелец. Патофизиология 16 (2-3): 233-46 (2009).
Епидемиологични доказателства за връзка между използването на безжични телефони и туморни заболявания. Hardell, Carlberg, et al. Патофизиология 16 (2-3): 113-122 (2009).
Употреба на мобилен телефон и риск от доброкачествени и злокачествени тумори на паротидната жлеза - общонационално проучване за контрол на случаи. Sadetzki, Chetrit, et al. Американски вестник по епидемиология 167 (4): 457-467 (2008).
V. Други злокачествени заболявания
Канцерогенният потенциал на нейонизиращите лъчения: Случаите на S-50 Hz MF и 1,8 GHz GSM радиочестотна радиация. Soffritti, M. и Giuliani, L. Основна и клинична фармакология и токсикология (2019).
Насърчаване на тумора чрез излагане на радиочестотни електромагнитни полета под границите на експозиция за хората. Lerchl, Klose, et al. Съобщения за биохимични и биофизични изследвания 459 (4): 585-590 (2015).
Шведският преглед укрепва основанията за заключение, че излъчването от клетъчни и безжични телефони е вероятен канцероген за човека. Davis, Kesari, et al. Патофизиология 20 (2): 123-129 (2013).
Мултифокален рак на гърдата при млади жени с продължителен контакт между гърдите им и техните клетъчни телефони. West, Kapoor, et al. Доклади в медицината (2013).
Епидемиологични доказателства за връзка между използването на безжични телефони и туморни заболявания. Hardell, Carlberg, et al. Патофизиология 16 (2-3): 113-122 (2009).
Изследване на потенциалните ефекти на "902 MHz GSM-тип безжични комуникационни сигнали" върху индуцирани от DMBA млечни тумори при плъхове Sprague-Dawley. Hruby, Neubauer, et al. Мутационни изследвания 649 (1-2): 34-44 (2008).
VI. Ефекти върху ДНК
Микровълните от мобилни телефони възпрепятстват формирането на фокус 53BP1 в човешките стволови клетки по-силно, отколкото в диференцираните клетки: Възможна механична връзка с риска от рак. Маркова, Малмгрен и др. Перспективи за здравето на околната среда 118 (3): 394-399 (2010).
Радиочестотна радиация и експресия на ген / протеин: Преглед. McNamee и Chauhan. Изследване на радиацията 172 (3): 265-287 (2009).
Оценка на експресията на HSP70 и увреждането на ДНК в клетките на човешка трофобластна клетъчна линия, изложени на амплитудно-модулирани радиочестотни полета с честота 1,8 GHz. Valbonesi, Franzellotto, et al. Изследване на радиацията 169 (3): 270-279 (2008).
Експресия на ген и протеин след излагане на радиочестотни полета от мобилни телефони. Vanderstraeten и Verschaeve. Перспективи за здравето на околната среда 116 (9): 1131-5 (2008).
Нетермални ефекти на експозицията на радиочестотно поле върху динамиката на калция в невронални клетки, получени от стволови клетки: изясняване на калциевите пътища. Rao, Titushkin, et al. Изследване на радиацията 169 (3): 319-329 (2008).
Промени в генната експресия в кожата на плъховете, предизвикани от продължително излагане на милиметрова вълна от 35 GHz. Millenbaugh, Roth, et al. Изследване на радиацията 169 (3): 288-300 (2008).
ДНК увреждане в Molt-4 Т-лимфобластоидни клетки, изложени на клетъчни телефонни радиочестотни полета in vitro. Philips, Ivaschuk, et al. Биоелектрохимия и биоенергетика 45 (1): 103-110 (1998).
VII. Неврологични / когнитивни ефекти
Ранното излагане на импулсни LTE радиочестотни полета причинява трайни промени в активността и поведението на мишки C57BL / 6 J. Broom, K., et al. Bio Electro Magnetics 40 (7): 498-511 (2019).
Повишаването на електромагнитното поле в човешката среда, взаимодействието с множество замърсявания на околната среда, ли е отправната точка за увеличаване на неврологичните смъртни случаи в Западния свят? Pritchard, C., Silk, A. и Hansen, L. Медицински хипотези 127: 76-83 (2019).
Ефект от 1800-2100 MHz електромагнитно излъчване върху учебната памет и хипокампалната морфология при швейцарски мишки албинос. Kishore, G., Venkatashu, K. и Sridevi, N. Jorunal от Clincal and Diagnostic Research 12 (2): 14-17 (2019).
Мониторинг на здравето на щам на мишки BALB / C, Изследване на поведението, хематологични параметри под въздействието на електромагнитно поле. Zymantiene, J., et al. Medycyna Weterynarjna 75 (03): 158-163 (2019).
2,45 GHz микровълнова радиация нарушава обучението, паметта и хипокампалната синаптична пластичност при плъховете. Karimi, N., et al. Токсикология и индустриално здраве 34 (12): 873-883 (2018).
Разстояние на мобилния телефон от главата и температурните промени на радиочестотните вълни в мозъчната тъкан. Forouharmajd, F., Ebrahimi, H. и Pourabdian, S. International Journal of Preventative Medicine (2018).
Проспективно кохортно проучване на ефективността на паметта на юношите и индивидуалната мозъчна доза на микровълнова радиация от безжична комуникация. Foerster, М., et al. Перспективи за здравето на околната среда 126 (7) (2018).
Електромагнитното излъчване 2450 MHz експозицията причинява дефицит на познание с митохондриална дисфункция и активиране на вътрешния път на апоптоза при плъхове. Gupta, SK, Mesharam, MK и Krishnamurthy, S. Journal of Biosciences 43 (2) 263-276 (2018).
Ефектът на Wi-Fi електромагнитните вълни при едномодални и мултимодални задачи за разпознаване на обекти при мъжки плъхове. Hassanshahi, A., et al. Неврологични науки 38 (6): 1069-1076 (2017).
Ефекти от краткосрочното и дългосрочно излагане на електромагнитни полета върху човешкия хипокампус. Deniz, OG, et al. Вестник по микроскопия и ултраструктура 5 (4): 191-197 (2017).
Ефекти от дългосрочното излагане на радиация 900-1800 MHz, излъчвана от 2G мобилен телефон върху мишки хипокампус - Истоморфометрично проучване. Mugunthan, Shanmugasamy, et al. Списание за клинични и диагностични изследвания 10 (8): AF01-6 (2016).
Ефект на излъчване на мобилен телефон върху праг на припадък, предизвикан от пентилентетразол при мишки. Kouchaki, Motaghedifard, et al. Ирански вестник на основните медицински науки 19 (7): 800-3 (2016).
Ефекти от 3 Hz и 60 Hz изключително нискочестотни електромагнитни полета върху подобно на тревожност поведение, запазване на паметта при пасивно избягване и електрофизиологични свойства на мъжките плъхове. Rostami, Shahani, et al. J Lasers Medical Science 7 (2): 120-125 (2016).
Краткосрочната памет при мишки се влияе от излъчването на мобилния телефон. Ntzouni, Stamatakis, et al. Патофизиология 18 (3): 193-199 (2011).
Използване на мобилни телефони и промени в когнитивните функции при юноши. Томас, Бенке и др. Медицина на труда по околна среда 67 (12): 861-866 (2010).
Повишена пропускливост на кръвно-мозъчната бариера в мозъка на бозайници 7 дни след излагане на радиация от мобилен телефон GSM-900. Nittby, Brun, et al. Патофизиология 16 (2-3): 103-112 (2009).
Ефекти на GSM 1800 MHz върху дендритното развитие на култивирани хипокампални неврони. Ning, Xu, et al. Acta Pharmacol Sin28 (12): 1873-1880 (2007).
Неврологични ефекти на радиочестотното електромагнитно излъчване. Лай, Хенри. Напредъкът в електромагнитните полета в живите системи1: 27-80 (1994).
VIII. Ефекти върху фертилитета при мъжете
Дългосрочното излагане на 4G смартфон радиочестотно електромагнитно излъчване намалява мъжкия репродуктивен потенциал чрез директно разрушаване на оста Spck3-MMP2-BTB в тестисите на възрастни плъхове. Yu, G., et al. Science of The Total Environment 698 (2020).
Радиации и мъжка плодовитост. Kesari, K., Agarwal, A. и Henkel, R. Репродуктивна биология и ендокринология 16 (118) (2018).
Ефектът от 2,45 GHz нейонизиращо лъчение върху структурата и ултраструктурата на тестисите при младите плъхове. Šimaiová, V., et al. Хистология и хистопатология 34 (4): 391-403 (2018).
Модулиращ ефект на радиацията 900 MHz върху биохимичните и репродуктивните параметри при плъхове. Narayana, SN., Et al. Братиславски медицински вестник 119 (9): 581-587 (2018).
Сокът от алое Arborescens предотвратява индуцирания от ЕМП оксидативен стрес и по този начин предпазва от патофизиология в мъжката репродуктивна система In Vitro. Solek, P., Majchrowics, L. и Koziorowski, M. Изследвания на околната среда 166: 141-149 (2018).
Радиочестотно излъчване (900 MHz), предизвикано от ДНК увреждане и арест на клетъчния цикъл в зародишните клетки на тестисите при швейцарски мишки албинос. Pandey, N., et al. Токсикология и индустриално здраве 33 (4) 373-384 (2017).
Ефектите на радиочестотното електромагнитно излъчване върху сперматозоидната функция. Хюстън, Никсън и др. Възпроизвеждане (2016)
Мъжката плодовитост и нейната връзка с професионалните опасности и опасностите от мобилни телефони: Аналитично проучване. Al-Quzwini, Al-Taee и др. Вестник на обществото за плодовитост в Близкия изток (2016).
Увреждане на ДНК на сперматозоиди - Ефектът от стреса и ежедневните фактори. Radwan, M, et al. Международен вестник за изследване на импотентността 28, 148-154 (2016).
Електромагнитното излъчване при 900 MHz предизвиква апоптоза на спермата чрез bcl-2, bax и каспаза-3 сигнални пътища при плъхове. Liu, Si, et al. Списание за репродуктивно здраве 12:65 (2015).
Навици при използване на мобилен телефон и качество на спермата - заслужава ли внимание? Zilverlight, Wiener-Megnazi, et al. Репродуктивна биомедицина онлайн 31 (3): 421-426 (2015).
Магнитните полета с изключително ниска честота предизвикват сперматогенна апоптоза на зародишните клетки: Възможен механизъм. Lee, Park, et al. BioMed Research International 2014 (2014): 567183 (2014).
In vitro ефект на радиацията на клетъчния телефон върху подвижността, фрагментацията на ДНК и експресията на клъстеринови гени в човешка сперма. Zalata, El-Samanoudy и др. Международен вестник за плодовитост и стерилност 9 (1): 129-136 (2014).
Ефект от излагането на електромагнитно поле върху репродуктивната система. Gye and Park. Списание за клинична и експериментална репродуктивна медицина 39 (1): 1-19 (2012).
Ефекти от експозицията на мобилни телефони върху мъжката репродукция: Преглед на литературата. Vignera, Condorelli, et al. Journal of Andrology 33 (3): 350-356 (2012).
Използването на преносими компютри, свързани към интернет чрез Wi-Fi, намалява подвижността на човешките сперматозоиди и увеличава фрагментацията на сперматозоидите. Avendano, C., et al. Плодовитост и стерилност 97 (1): 39045 (2012).
Излагане на магнитни полета и риск от лошо качество на спермата. Li, Yan, et al. Списание за репродуктивна токсикология 29 (1): 86-92 (2010).
Излъчването на мобилен телефон предизвиква производство на реактивен кислород и увреждане на ДНК при човешки сперматозоиди In Vitro. Luliis, Newey, et al. PLoS ONE 4 (7) (2009).
Радиочестотното електромагнитно излъчване (Rf-EMR) от GSM мобилни телефони предизвиква оксидативен стрес и намалява подвижността на сперматозоидите при плъхове. Mailankot, Kunnath, et al. Clinical Science 64 (6): 561-5 (2009).
Мобилни телефони: Немезида на съвременния човек? Makker, Varghese, et al. Репродуктивна биомедицина онлайн 18 (1): 148-157 (2008).
Ориентировъчни нива на SAR поради активен мобилен телефон в джоба на предния панталон в близост до общи метални предмети. Whittow, Panagamuwa, et al. Конференция за размножаване 149-152 (2008).
Клетъчни телефони и мъжко безплодие: дисекция на връзката. Deepinder, Makker, et al. Репродуктивна BioMedicine Online 15 (3): 266-270 (2007).
Оценка на ефекта от използването на мобилни телефони върху мъжката плодовитост. Wdowiak, Wiktor, et al. Annals of Agricultural and Medicine14 (1): 169-172 (2007).
Ефект от използването на мобилен телефон върху анализа на спермата при мъже, посещаващи клиника по безплодие: Наблюдателно проучване. Agarwal, Deepinder, et al. Американско общество за репродуктивна медицина 89 (1): 124-8 (2008).
IX. Електромагнитна чувствителност
Ставане на електро-свръхчувствителност: Проучване на репликацията. Dieudonne, M. Биоелектромагнитни 40: 188-200 (2019).
Функционален мозъчен ЯМР при пациенти, оплакващи се от електрохиперчувствителност след продължително излагане на електромагнитни полета. Heuser, G. & Heuser, S. Рецензии за здравето на околната среда 32 (3): 291-299 (2016).
Hot Nano Spots "като интерпретация на така наречените нетермични биологични ефекти върху мобилния телефон. Pfutzner, Helmut. Journal of Electromagnetic Analysis and Applications 8 (3): 62-69 (2016).
Анализ на генотоксичните ефекти на излъчването на мобилен телефон с помощта на анализ на букална микроядра: сравнителна оценка. Banerjee, Singh, et al. Вестник за клинични и диагностични изследвания 10 (3): ZC82-ZC85 (2016).
Шум в ушите и клетъчни телефони: Ролята на електромагнитното радиочестотно излъчване. Медейрос и Санчес. Бразилски вестник по оториноларингология 82 (1): 97-104 (2016).
Микровълновите честотни електромагнитни полета (ЕМП) произвеждат широко разпространени невропсихиатрични ефекти, включително депресия. Pall, Martin L. Journal of Chemical Neuroanatomy (2015).
Субективни симптоми, свързани с излъчването на GSM от базови станции за мобилни телефони: Проучване в напречно сечение. Gomez-Perretta, Navarro, et al. BMJ Open 3.12 (2013).
Зелена комуникация - Условие за намаляване на електромагнитната свръхчувствителност от клетъчни телефони. Kumar, Khan, et al. Procedia Technology 4: 682-686 (2012).
Електромагнитна свръхчувствителност: факт или измислица? Гений и Лип. Science of the Total Environment 414 (1): 103-112 (2012).
Радиочестотна (RF) болест в проучването Lilienfeld: Ефект от модулирани микровълни? Liakouris, A. Архиви на здравето на околната среда 236-238 (2010).
Невроповеденчески ефекти сред обитателите около базовите станции на мобилни телефони. Abdel-Rassoul, El-Fateh, et al. NeuroToxicology28 (2): 434-440 (2007).
Електро свръхчувствителност: Съвременно състояние на функционално увреждане. Йохансон, О. Електромагнитна биология и медицина 25 (4): 245-258 (2006).
Електромагнитна свръхчувствителност: биологични ефекти от мръсното електричество с акцент върху диабета и множествената склероза. Havas, M. Електромагнитна биология и медицина 25 (4): 259-268 (2006).
Установяването на здравни рискове от излагане на радиочестотни полета изисква мултидисциплинарни изследвания. Хиетанен, Майла. Скандинавски вестник за труда, околната среда и здравето 32 (3): 169-170 (2006).
Свръхчувствителност на човешките субекти към излагане на електрическо и магнитно поле в околната среда: Преглед на литературата. Левалоа, Патрик. Перспективи за здравето на околната среда 110 (4): 613-8 (2002).
Електрическа свръхчувствителност и неврофизични ефекти на клетъчните телефони - факти или излишно безпокойство? Харма, Мико Илмари. Скандинавски вестник за труда, околната среда и здравето 26 (2): 85-86 (2000).
X. Ефекти върху имплантираните медицински изделия
Ad Hoc изпитване за електромагнитна съвместимост на неимплантируеми медицински изделия и радиочестотна идентификация. Сейдман и Гуаг. Биомедицинско инженерство OnLine 12:71 (2013).
Електромагнитни смущения на пейсмейкърите. Lakshmanadoss, Chinnachamy, et al. Interchopen 229-252 (2011).
Намеса между мобилни телефони и пейсмейкъри: Поглед отвътре. Censi, Calcagnini, et al. Annali dell'Istituto superiore di sanità 43 (3): 254-259 (2007).
Електромагнитни смущения на пейсмейкърите. Ердоган, Окан. Indian Pacing and Electrophysiology Journal 2 (3): 74-78 (2002).
Електромагнитни смущения при пациенти с имплантирани кардиовертер-дефибрилатори и имплантируеми рекордери. Sousa, Klein, et al. Indian Pacing and Electrophysiology Journal 2 (3): 79-84 (2002).
Радиочестотни смущения с медицински изделия. Изявление за техническа информация. IEEE комисия по човека и радиацията, Институт на инженерите по електротехника и електроника 17 (3): 111-4 (1998).
Клетъчни телефони и пейсмейкъри: Спешно обаждане или грешен номер? Еленбоген и Ууд. Вестник на Американския колеж по кардиология 27 (6): 1478-9 (1996).
XI. 5G ефекти
Модел на стационарно повишаване на температурата в многослойни тъкани поради излагане на радиочестотно поле с милиметрови вълни с тесни лъчи. Gajda, G., et al. Физика на здравето 117 (3): 254-266 (2019).
Нецелена метаболомика разкрива промени на пропускливостта на биомембраните в HumanHaCaT кератиноцити при експозиция от 60 HGz милиметрова вълна . Pogam, Pierre., Et al. Научни доклади 9 (9343) (2019).
Очна реакция на излагане на милиметрови вълни при различни нива на влажност. Kojima, М., et al. Journal of Infrared Milli Terahz Waves 40: 574–584 (2019).
Излъчването на милиметрова вълна активира носицептори на пиявици чрез TRPV1-подобна сенсибилизация на рецептора. Romanenko, S., et al. Biophysical Journal 116 (12): 2331-2345 (2019).
Систематично извеждане на граници на безопасност за варираща във времето 5G радиочестотна експозиция въз основа на аналитични модели и термична доза. Neufeld, E. и Kuster, N. Health Physics Society (2018).
Към 5G комуникационни системи: Има ли последици за здравето? Ciaula, AD. Международен вестник по хигиена и здраве на околната среда 367-375 (2018).
Разширяване на 5G безжични телекомуникации: последици за общественото здраве и околната среда. Ръсел, CL Изследване на околната среда 165: 484-495 (2018).
Човешката кожа като приемник под THZ - представлява ли 5G опасност за нея или не? Betzalel, N., Ishai, PB и Feldman, Y. Изследвания на околната среда163: 208-216 (2018).
Моделиране на абсорбцията на слънчево-THz лъчение от човешка кожа. Betzalel, N., Feldman, Y. и Ishai, PB IEEE транзакции на Terahertz Science and Technology 7 (5): 521-528 (2017).
Излагане на хора на радиочестотни полета в 5G Downlink. Насим, И. и Ким, Южен университет на Джорджия (2017).
Човешкото тяло и безжичните комуникационни системи с милиметрови вълни: Взаимодействия и последици. Wu, T., Rappaport, T. и Collins, C. Международна конференция по комуникации IEEE (2015).
Състояние на знанията за биологичните ефекти при 40-60 GHz. Drean, Y., et al. Comptes Rendus Physique (2013).
Ефекти от излъчването на милиметрови вълни върху клетъчната мембрана-Кратък преглед. Рамундо-Орландо, Алфонсина. Вестник на инфрачервените, милиметровите и терагерцовите вълни 31 (12): 1400-1411 (2010)
Човешката кожа като масиви от спираловидни антени в милиметрови и субмилиметрови вълни. Feldman, Y., et al. Американското физическо общество (2008).
XII. Разни статии
Нецелена метаболомика разкрива промени в пропускливостта на биомембраните в човешки HaCaT кератиноцити при експозиция от 60 HGz милиметрова вълна. Pogam, Pierre., Et al. Научни доклади 9 (9343) (2019).
Рискове за здравето и благосъстоянието от радиочестотното излъчване, излъчвано от клетъчни телефони и други безжични устройства. Miller, A., et al. Frontiers in Public Health 7 (223) (2019).
Компютърна Simula ции о е проникването на 0,30 THz Радиация в човешкото ухо. Vilaagosh, Z., et al. Biomedical Optics Express 10 (3) (2019).
Излагане на радиочестотно електромагнитно поле и възприемане на риска: Пилотно експериментално проучване. Zeleke, B., et al. Екологични изследвания 170: 493-499 (2019).
Коментар за полезността на изследването на Националната токсикологична програма за радиочестотни данни за радиочестотна радиация за оценка на рисковете за човешкото здраве въпреки необоснованите критики, насочени към минимизиране на констатациите за неблагоприятни ефекти върху здравето. Melnick, R. Изследване на околната среда 168: 1-6 (2019).
Патологични находки, наблюдавани в бъбреците на постнатални мъжки плъхове, изложени на 2100 MHz електромагнитно поле. Bedir, R., et al. Архиви на медицински изследвания (2019).
Генотоксични и канцерогенни ефекти на нейонизиращите електромагнитни полета. Kocaman, A., et al. Екологични изследвания 163: 71-79 (2018).
Нейонизираща опасност от ЕМП през 21 век . Koh, WJ и Moochhala, SM IEEE (2018).
Топлинни и нетермични здравни ефекти на нейонизиращото лъчение с ниска интензивност: международна перспектива. Belpomme, D., et al. Замърсяване на околната среда 242 (A): 643-658 (2018).
Сравнение на нивата на експозиция на радиочестотното електромагнитно поле в различни ежедневни микросреди в международен контекст. Sagar, S, et al. Environmental International 114: 297-306 (2018).
Wi-Fi е важна заплаха за човешкото здраве. Пал, М. Изследвания на околната среда 405-416 (2018).
Възбуждане, предизвикано от радиация на мобилния телефон, на профилиране на генно-експресионно равновесие и контрол на спорадична апоптоза в яйчника на Drosophila Melanogaster. Manta, A., et al. FLY 11 (2): 75-95 (2017).
Световна здравна организация, Радиочестотна радиация и здраве - твърд орех, който трябва да се напука (Преглед). Hardell, L. International Journal of Oncology51: 405-413 (2017).
Излъчването от безжична технология повишава кръвната глюкоза и телесната температура при 40-годишен мъж с диабет тип 1. Kleiber, C. Електромагнитна биология и медицина 36: 3 259-264 (2017).
Сърдечно-съдови заболявания: Време е да се идентифицират възникващите фактори на околната среда. Bandara, P. & Weller, S. Европейско списание за превантивна кардиология (2017).
Ефекти от експозицията на 2100MHz GSM-подобно радиочестотно електромагнитно поле върху слухова система на плъхове. Celiker, Ozgur, et al. Бразилски вестник по оториноларингология (2017).
Изследване на ефекта от изключително нискочестотни импулсни електромагнитни полета върху човешки електрокардиограми (ЕКГ). Fang, Mahmoud, et al. Международен вестник за изследване на околната среда и обществено здраве 13 (11) (2016).
Оценка на защитната роля на витамин С върху метаболитните и енксиматични дейности на черния дроб при мъжките плъхове след излагане на 2,45 GHz Wi-Fi рутери. Shekoohi-Shooli, F., et al. Вестник по биомедицинска физика и инженерство 6 (3): 157-164 (2016).
Излагането на ELF-EMF и RF-EMF увеличава скоростта на транспортиране на глюкоза и TCA цикъл в пъпки дрожди. Lin, Yan, et al. Frontiers in Microbiology (2016).
Кампания за повишаване на осведомеността срещу радиационната опасност от мобилен телефон: казус Оман. Осмен и Саар. Procedia - Социални и поведенчески науки 205 (9): 381-385 (2015).
Електромагнитната енергия, излъчвана от мобилния телефон, променя електрокардиографските записи на пациенти с исхемична сърдечна болест. Alhusseiny, Al-Nimer, et al. Annals of Medical and Health Science Research 2 (2): 146-151 (2012).
Ефекти на радиочестотното излъчване върху човешкия феритин: In vitro анализ на ензима. Fattahi-asl, Baradaran-Ghahfarokhi, et al. Journal of Medical Signals and Sensors 2 (4): 235-240 (2012).
Апоптозата се индуцира от радиочестотни полета през независимия от каспаза митохондриален път в кортикалните неврони. Joubert, Bourthoumieu, et al. Изследване на радиацията 169 (1): 38-45 (2008).
Източник на финансиране и резултати от изследвания на здравните ефекти от използването на мобилни телефони: Систематичен преглед на експериментални изследвания. Huss, Egger, et al. Перспективи за здравето на околната среда 115 (1): 1-4 (2007).
Епидемиология на здравните ефекти от радиочестотната експозиция. Ahlbom, Green, et al. Перспективи за здравето на околната среда 112 (17): 1741-1753 (2004).
Възможната роля на радиочестотното излъчване в развитието на Uveal меланом Stang, Anastassiou и др. Journal of Epidemiology 12 (1): 7-12 (2001).
Биологични ефекти от амплитудно модулирана радиочестотна радиация. Юутилайнен и Сезе. Скандинавски вестник за труда, околната среда и здравето 24 (2): 245-254 (1998).


Коментари
Публикуване на коментар